Can viruses provide key information on the evolution of cells?
Published in Microbiology

Although, according to Yogi Berra, predictions about the future are particularly challenging, retrodiction is not much easier when it comes to the distant past. Thus, the origin of the first cells, the nature of the Last Universal Cellular Ancestor (LUCA), the paths from the LUCA to bacteria and archaea, the two primary domains of life, and the origin of eukaryotes (eukaryogenesis) are among the hardest as well as the most fascinating problems in biology.
In recent years, the study of eukaryogenesis made a major leap forward thanks to the discovery and exploration of a new phylum of archaea that received the picturesque name Asgard, after the mythical abode of the Nordic gods (all the names of archaeal taxa within Asgard come from particular deities, the first one to be discovered being the mischievous Loki). In phylogenetic trees of universally conserved genes, primarily encoding translation system components, eukaryotes unequivocally fall within Asgard and seem to show the closest affinity with the Asgard class Heimdallarchaeia (after Heimdall, the guardian god). No less importantly, genomic analysis of Asgard archaea revealed homologs of many genes that were previously considered eukaryotic signatures, especially, those encoding components of the cytoskeleton and intracellular membrane remodeling systems. There is now effectively no doubt that the archaeal parent of eukaryotes was an Asgard archaeon. Was this archaeon the host of the alphaproteobacterial endosymbiont that became the ancestor of the mitochondria, giving rise to the first eukaryotic cells? Perhaps… but the most plausible scenario of eukaryogenesis actually might be not as simple as that.
The picture of life evolution cannot be complete without understanding evolution of viruses
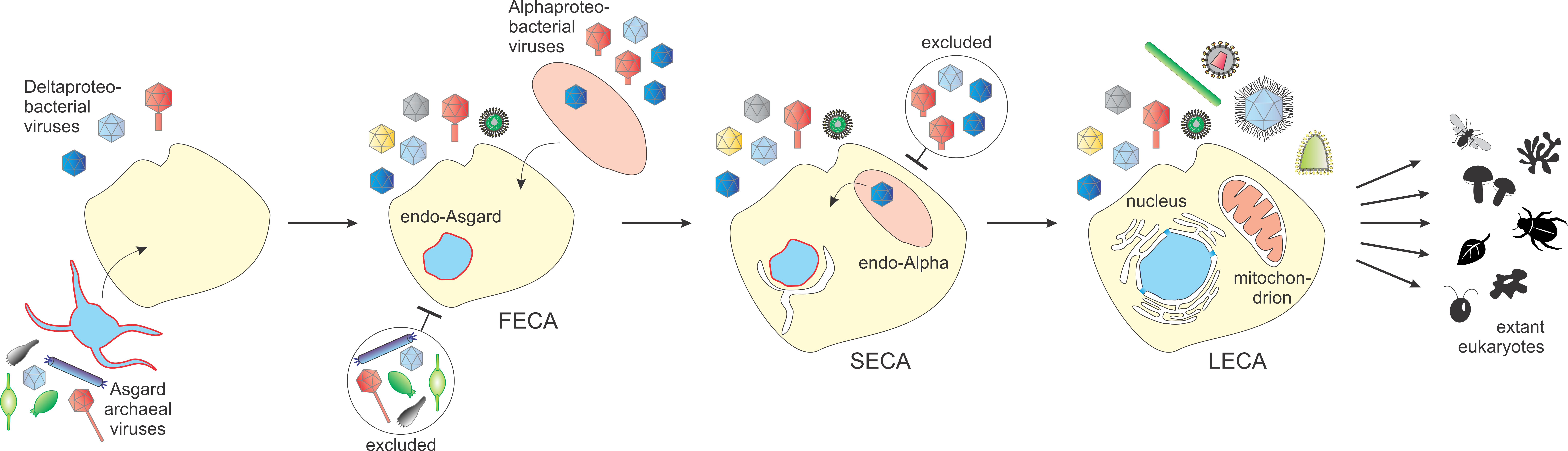
All cellular life forms (with a few explainable exceptions) are hosts to multiple viruses. There is every reason to believe that viruses accompanied life since the earliest stages of its evolution, emerging soon after the first cells. Therefore, reconstruction of ancestral life forms can hardly be considered complete without an accompanying reconstruction of the entirety of viruses that infected them, the virome, for short. In the article we just published in Nature Microbiology with Mart Krupovic and Valerian Dolja, we undertake the reconstruction of the virome of the Last Eukaryotic Common Ancestor (LECA). This is the latest in our series of efforts to address the fundamental problems in the evolution of viruses, the previous installments dealing with the origin of viruses, the virome of the LUCA and the logic of virus evolution, that is, what types of genes viruses capture from the hosts and how these genes are adapted for viral functions. The approach used for the reconstruction of ancestral viromes is simple and straightforward: we map different groups of viruses on the phylogenetic tree of the hosts and construct what is called the most parsimonious scenario of evolution, that is, the shortest sequence of events (in this case, gain or loss of a given virus group) that could generate the observed distribution. The results of such reconstruction for the LUCA and the LECA recapitulated each other. In both cases, we found that the ancestral viromes were already highly complex and diverse including representatives from most of the major groups of extant viruses.
This result is best compatible with rapid, explosive diversification of viruses both at the early stage of evolution antedating the LUCA and during eukaryogenesis. Not unexpectedly, the reconstructed virome of LUCA resembled today’s prokaryote viromes, whereas the virome of LECA included the main groups of viruses of eukaryotes. The viromes of eukaryotes and prokaryotes are dramatically different, but nevertheless, the prokaryotic roots of the majority of eukaryotic viruses can be traced by comparison of viral genomes and virion structures. And here came a surprise: we realized that all main types of viruses of eukaryotes apparently originated from viruses infecting bacteria (bacteriophages) rather than archaeal viruses. At least superficially, this is unexpected because viruses are genetic, informational parasites, and in eukaryotes, the information processing systems are predominantly of archaeal origin. And then, there was a more concrete reason why the bacterial provenance of the LECA virome surprised us. Three recent studies published in Nature Microbiology, from the laboratories of Thijs Ettema, Mart Krupovic and Brett Baker analyzed for the first time viruses of Asgard archaea. Some of these viruses were found to be related to those of other archaea whereas others appeared largely unique, but there was no detectable evolutionary connections at all with viruses of eukaryotes. There is at this point no reasonable doubt that an Asgard archaeon was one of the partners in the symbiosis that gave rise to eukaryotes and, certainly, that archaeon was infected by a number of different viruses. Why then were these viruses banished from the emerging eukaryotes and replaced with the bacterial viruses?
Eukaryogenesis: one or two symbioses?
The moment of truth came in a rather casual discussion one of us (E.V.K) had with a colleague who made important contributions to our understanding of eukaryogenesis (more about that below), Puri Lopez-Garcia of Université Paris-Saclay/CNRS. To the above question on the bacterial provenance of virus infecting eukaryotes, Puri dropped casually: Perhaps, it is because of the membranes? And that was it, so embarrassingly simple: certainly, it is all about the membranes! One of the major differences between bacterial and archaeal cells is the different chemical structure of their membranes which are built of unrelated phospholipids. Crucially, all membranes in eukaryotic cells are of the bacterial type. For any model of eukaryogenesis, this difference between bacterial and eukaryotic membranes, on the one hand, and archaeal membranes on the other hand, is a major challenge. The simplest symbiogenetic scenarios, which involve an archaeal host (now shown to come from the Asgard phylum) and an alphaproteobacterial proto-mitochondrial endosymbiont as the only two partners in eukaryogenesis, face the problem of replacing the archaeal membrane with the bacterial one, for which we know no precedent. The bacterial provenance of the LECA virome made us to turn to alternative eukaryogenesis models, developed by Puri Lopez-Garcia with her longtime collaborator David Moreira, in which the emerging protoeukaryote never had an archaeal cell membrane at all. These are known as syntrophy models because they were initially motivated by the plausibility of a metabolic symbiosis between a hydrogen-producing deltaproteobacterium and a methanogenic archaeon. These models postulate two consecutive symbioses, where the deltaproteobacterium engulfed the archaeon at the first stage, and the resulting chimeric organism subsequently captured the alphaproteobacterium that became the mitochondrion (see Figure below). Admittedly, the requirement for two endosymbiotic events instead of one adds complexity to the eukaryogenesis scenario, and probably, for that reason, the original syntrophy model did not seem to win over the students of eukaryogenesis. However, the recent demonstration of the syntrophic relationships between Asgard archaea and deltaproteobacteria seems to be a game changer, an important if indirect support of a syntrophy scenario for eukaryogenesis.
In the current version of the syntrophy model, eukaryogenesis started as a metabolic ectosymbiosis between a sulfate-reducing deltaproteobacterium and a hydrogen-producing Asgard archaeon. The archaeon was subsequently internalized and lost its membrane, likely, after the emergence of bacterial endomembranes that surrounded the engulfed archaeon. The internalized archaeon became the progenitor of the eukaryotic nucleus. The merger between a deltaproteobacterium host and an archaeal endosymbiont gave rise to an evolutionary intermediate, First Eukaryotic Common Ancestor (FECA). This first endosymbiotic event was followed by the secondary endosymbiosis, whereby FECA gave rise to Second Eukaryotic Common Ancestor (SECA) by capturing a versatile sulfur-oxidizing and facultatively aerobic alphaproteobacterium that became the mitochondrion of the LECA. The two endosymbiotic events in this scenario might not be as big a deal as it seems. Indeed, serial endosymbiosis in the subsequent evolution of eukaryotes was rather common, evolution of chloroplasts from cyanobacteria in the ancestor of red and green algae and plants being only one, best known example. Furthermore, this scenario is compatible with independent phylogenomic analysis indicating that mitochondrial endosymbiosis occurred relatively late in the evolution of eukaryotes. Add to this that enzymes of certain metabolic pathways in eukaryotes do seem to be of deltaproteobacterial origin.
Crucially, under the syntrophy scenario, the emerging eukaryotic cell maintained the bacterial membranes through all stages of eukaryogenesis, whereas the archaeal membrane of the primary endosymbiont was lost. This continuity of bacterial membranes throughout eukaryogenesis seems to solve the problem of the bacterial provenance of the eukaryotic virome. Indeed, it is highly likely that viruses of the Asgard archaeon were excluded following its internalization during the first stage of eukaryogenesis, primarily, due to the inaccessibility of the archaeal virus receptors. Arguably, the escape from viruses facilitated the endogenization of the archaeal symbiont en route to FECA, jumpstarting eukaryogenesis. Under this scenario, the LECA virome was shaped by two waves of adaptation of bacterial viruses, first, from the deltaproteobacterial virome, and then, from the virome of the alphaproteobacterial proto-mitochondrion (see Figure).
Coming back to where we started, reconstructing singular events, such as eukaryogenesis, that led to major evolutionary transitions in the deep past (in this case, 1.5-2 billion years ago) is a formidable task. And yet, the problem does not appear unassailable. The discovery of Asgard archaea with their close relationship with eukaryotes and the syntrophic lifestyle is obvious progress. The bacterial provenance of the eukaryote virome further buttresses models of eukaryogenesis that postulate the evolutionary continuity of bacterial membranes, such as the syntrophy scenario. It seems reasonable to hope that future research into the biology of Asgard archaea and their interactions with bacteria as well as detailed characterization of the viromes of all partners involved in eukaryogenesis helps us understand this pivotal event much better. We might never know how it happened, exactly, but specific models testable in the laboratory could be a realistic goal.
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in