The COVID-19 pandemic, year 2: Bitter lessons in evolution and ecology
Published in Microbiology
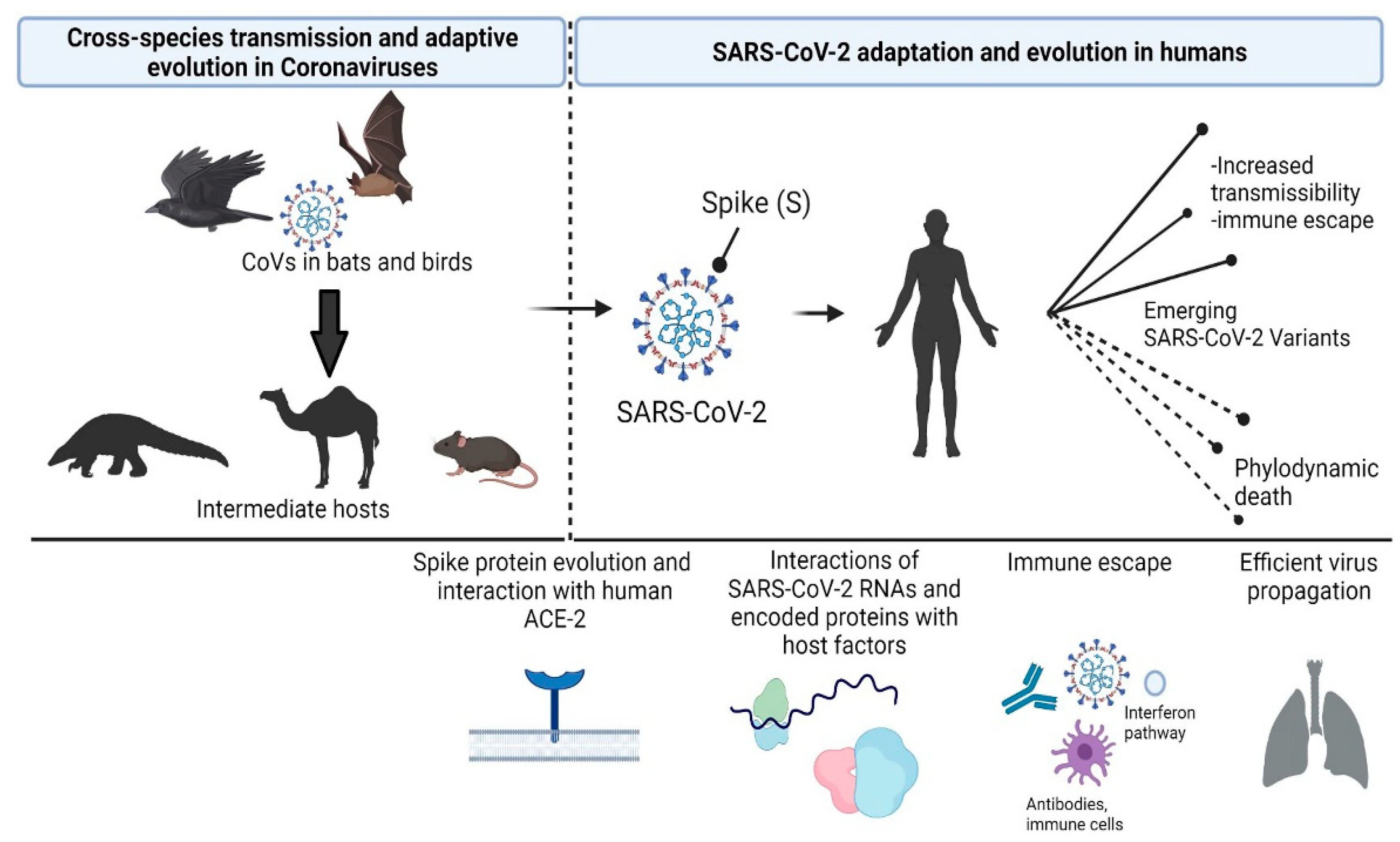
As we step on the threshold of 2022, the coronavirus pandemic that originated in Wuhan, China during December 2019 continues to cast a long shadow on the future. As a pathogen that has recently 'jumped' from an animal host (bats) to humans, it is quite likely that SARS-CoV-2 will take its time before becoming endemic and seasonal across large regions of the globe1–3. En route, it has amply highlighted the signal importance of ecological and evolutionary considerations in understanding the course of infectious diseases.
The outbreak became a pandemic in great part due to the enhanced and rapid means of mobility that industrial society has placed at our disposal. It is interesting to note that the possibility of a pandemic due to the availability of rapid means of worldwide travel was noted by Nassim Taleb in his book The Black Swan4 in 2007. The historically unprecedented connectivity of the world has introduced an important ecological factor into the dynamics of infectious diseases in general and COVID-19 in particular5. Besides, SARS-CoV-2 has proven its capability to utilize a slew of opportunities for its spread and has made us conscious of the very definition of ecology - the study of organisms and their relationships with their habitat. From the viral viewpoint, we happen to be its most recent “habitat” that it has stumbled upon. It has won the ecological jackpot in its infection of a host (habitat) that is sufficiently numerous (unlike say, the blue whale), is present on every continent except Antarctica (unlike penguins or apes), is markedly gregarious (unlike for example, the sloth or the tiger) and can travel between any two far-flung regions of the globe almost at will (unlike migratory birds that tend to follow well-traveled routes between traditional habitats and only at specified times of the year). No other large animal would satisfy ALL these criteria. This is not to be construed that SARS-CoV-2 itself is seeking an ideal or an optimal host; it seems quite capable of infecting pets6 and wild animals7 alike in a case of reverse zoonosis. Rather, the ability of the spike protein to bind to the ACE2 receptor whose homologs are shared among many mammals8–10 is likely what enables it to infect multiple hosts. Thankfully, in most cases, the disease itself is relatively mild and individuals recover satisfactorily. This is a textbook case of the ecological principle that a predator must not be so efficient as to completely exterminate its prey, only to suffer extinction itself in consequence. Given that the upper respiratory tract is the virus’ primary habitat and portal of infection means that it can be efficiently transmitted by means of respiratory droplets.
Testing for infection in large numbers and in real time has demonstrated that the incidence of infection in different populations waxes and wanes with time – hence the talk about ‘waves’ of infections. As an RNA virus capable of replicating to large numbers in a very short time, besides harbouring a more error-prone replication machinery, SARS-CoV-2 has perforce provided us a ringside view of evolution. Not that this is unique to SARS-CoV-2; the influenza vaccine is routinely modified every year based on the particular variants that are expected to dominate the viral population in a given year. As the adaptive immune system of infected (or vaccinated) individuals imposes selection pressure on the rapidly multiplying viral population, new variants capable of escaping neutralizing antibodies come to dominate the next round of infections. The omicron variant that seems to be spreading rapidly now is one visible consequence of this fact11. One problem with this particular virus is not just that it is an additional pathogen among many; rather, it is what is termed an 'emerging,' or more appropriately, a 'newly emergent' pathogen and has not co-evolved with its new host. Bats, by contrast, seem to have adapted to this (and other) viruses by having balanced immune tolerance and defense12 and seem to avoid the serious and sometimes lethal complications of infection. And when it comes to variation, all human beings are not cut from the same (genetic) cloth either13,14. Among those who survive infection, we have asymptomatic carriers on one extreme, and those that suffer serious consequences of COVID-19 for an extended period of time after the virus itself has been cleared – the COVID long haulers – on the other15. The damage response framework could be an appropriate theoretical lens through which we may understand the varying clinical outcomes of COVID-19 in different individuals16,17.
So, the way forward seems arduous, to say the least but not entirely impassable or worse, trackless. If viral transmission can be reduced by orders of magnitude by the simple expedient of masking18 and elementary hygienic practices in addition to vaccination, there is hope that this virus can be contained in the future. While we cannot really do anything about viral virtuosity in terms of its genetics or the genetic hand that we were dealt at conception, we can make a difference in terms of the ecological opportunities we present to the virus by means of our behaviour. In this situation, one cannot help but remember the late Victor Frankl (Holocaust survivor and the founder of logotherapy) who wrote as follows19:
“In fact, freedom is in danger of degenerating into mere arbitrariness unless it is lived in terms of responsibleness. That is why I recommend that the Statue of Liberty on the East Coast be supplemented by a Statue of Responsibility on the West Coast.”
Every infection prevented is an invisible but nevertheless tangible gain for individuals and healthcare systems alike.
Notes:
Poster image from Fig. 1 in Beloukas & Rampias (2021)20 under the Creative Commons Attribution License 4.0 (CC BY).
Disclaimer: The above post is not intended as medical or policy advice. No guarantee is expressed or implied regarding the veracity and medical utility of the information provided on external websites and sources. The opinions expressed herein do not represent the views of the TERI School of Advanced Studies or TERI.
References:
- Eguia, R. T. et al. A human coronavirus evolves antigenically to escape antibody immunity. PLOS Pathog. 17, e1009453 (2021).
- Callaway, E. Beyond Omicron: what’s next for COVID’s viral evolution. Nature 600, 204–207 (2021).
- Murray, C. J. L. & Piot, P. The Potential Future of the COVID-19 Pandemic: Will SARS-CoV-2 Become a Recurrent Seasonal Infection? JAMA 325, 1249–1250 (2021).
- Taleb, N. N. The Black Swan: the impact of the highly improbable. (Penguin Books, 2010).
- Cirillo, P. & Taleb, N. N. Tail risk of contagious diseases. Nat. Phys. 16, 606–613 (2020).
- Sit, T. H. C. et al. Infection of dogs with SARS-CoV-2. Nature 586, 776–778 (2020).
- Chandler, J. C. et al. SARS-CoV-2 exposure in wild white-tailed deer (Odocoileus virginianus ). Proc. Natl. Acad. Sci. 118, e2114828118 (2021).
- Cao, Y. et al. Analysis of ACE2 Gene-Encoded Proteins Across Mammalian Species. Front. Vet. Sci. 7, (2020).
- Damas, J. et al. Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc. Natl. Acad. Sci. 117, 22311–22322 (2020).
- Wei, Y., Aris, P., Farookhi, H. & Xia, X. Predicting mammalian species at risk of being infected by SARS-CoV-2 from an ACE2 perspective. Sci. Rep. 11, (2021).
- Liu, L. et al. Striking antibody evasion manifested by the Omicron variant of SARS-CoV-2. Nature (2021) doi:10.1038/d41586-021-03826-3.
- Irving, A. T., Ahn, M., Goh, G., Anderson, D. E. & Wang, L.-F. Lessons from the host defences of bats, a unique viral reservoir. Nature 589, 363–370 (2021).
- COVID-19 Host Genetics Initiative et al. Mapping the human genetic architecture of COVID-19. Nature 600, 472–477 (2021).
- Colona, V. L., Vasiliou, V., Watt, J., Novelli, G. & Reichardt, J. K. V. Update on human genetic susceptibility to COVID-19: susceptibility to virus and response. Hum. Genomics 15, (2021).
- Rajkumar Tripathi, A. K. & Pinto, L. M. Long COVID: ‘And the fire rages on’. Lung India Off. Organ Indian Chest Soc. 38, 564–570 (2021).
- Pirofski, L.-A. & Casadevall, A. Pathogenesis of COVID-19 from the Perspective of the Damage-Response Framework. mBio 11, (2020).
- Casadevall, A. & Pirofski, L. The damage-response framework of microbial pathogenesis. Nat. Rev. Microbiol. 1, 17–24 (2003).
- Kollepara, P. K., Siegenfeld, A. F., Taleb, N. N. & Bar-Yam, Y. Unmasking the mask studies: why the effectiveness of surgical masks in preventing respiratory infections has been underestimated. J. Travel Med. 28, (2021).
- Frankl, V. E., Lasch, I. & Kushner, H. S. Man’s search for meaning: the classic tribute to hope from holocaust. (Rider, 2008).
- Beloukas, A. & Rampias, T. Biological and Clinical Significance of Adaptive Evolution of Coronaviruses. Life 11, 1129 (2021).




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in