A chromosome in the trap is worth two in the bush
Published in Physics
When eukaryotic cells are dividing, they need to make sure that each newly formed daughter cell inherits one copy of the duplicated genome. What sounds straightforward is actually an incredibly challenging task. During interphase (the time in the cell cycle between cell division), the DNA of eukaryotic cells, which is stored in the nucleus, needs to be accessible for the transcription machinery reading the genetic code. That means that in this time of the cell cycle the DNA is loosely packed, so that the necessary proteins for transcription can bind to the DNA and move along it. Anybody who’ve had a pair of headphones in their pocket for a while, pulls them out and tries to disentangle them, can understand the difficulty of separating these intermingled chromosomes. This alone would be challenging enough, but gets further complicated by the astonishing amounts of DNA stored in the nucleus. A human cell has around 2 m of DNA, that is all packed in the nucleus of only 10 µm size. In order to tackle this challenge, eukaryotic cells use a trick: They form the DNA into a very compact and defined structure, the mitotic chromosome. This helps in two ways: on the one hand this compaction separates different chromosomes from each other, but it also allows to easily align them in the mitotic plane between the two nascent daughter cells, such that each new cell ends up with exactly one copy of each chromosome.
 Diagram of the transitions from interphase through the stages of mitosis, (right) entangled headphones.")
Although biologists have studied mitotic chromosomes for roughly 150 years using microscopy and biochemical methods, we still do not fully understand their internal structure. This is particularly due to their very high density and the tight packing of DNA and proteins in the chromosome, making it incredibly hard to take a peek inside of a mitotic chromosome. In addition, most studies have been on chemically fixed chromosomes. It is a bit like studying wild animals, but you only have access to stuffed specimens. In our recent paper “Nonlinear mechanics of mitotic chromosomes”, we and our co-authors therefore describe a new way of studying native mitotic chromosomes in great detail using optical tweezers to get insights into their internal organization.
It all started with an idea five years ago. If we can do detailed optical tweezer analysis of DNA molecules, why not chromosomes? This led to the initial awkward meetings between physicists from the Peterman/Wuite lab and biologists from the Hickson lab. Because, “how do you speak to physicists AND understand what they are saying?”, to quote Ian Hickson. Along the way physicists had to learn biology and biologists had to learn physics. Step by step, this (as well as a few beers) allowed us in the end to speak the same language. “It’s been a truly exciting experience with many lessons learned, mistakes made and challenges overcome” as Gijs Wuite summarized it.
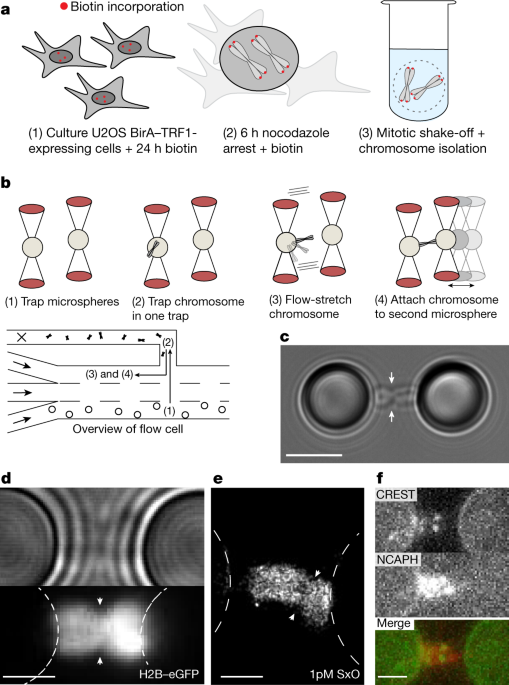
The first challenge was how to prepare chromosomes for optical tweezer analysis, while still retaining their native structure. First, we needed some sort of molecular handle, to grab the chromosome: We genetically engineered virus particles, which upon infection of cells would attach biotin handles to the telomeric ends of chromosomes. These biotin molecules can form a tight bond with the protein streptavidin, which we use to attach these chromosomes to micrometer-sized polystyrene beads, which then in turn are held by our optical tweezers. We then developed a method to isolate chromosomes in a way that preserves their native structure without chemical fixation. Our colleague Kata Sarlos remembers, “Christian and I went on an excursion to the William Earnshaw lab in Edinburgh to learn from the best, this improved our chromosome isolation protocol greatly”. These isolated chromosomes are suitable to be caught with optical tweezers, highly focused laser beams capable of trapping and moving small objects. Being able to catch a chromosome was the major breakthrough in this project, as our colleague Anna Meijering recalls: “I still remember the first time I trapped an actual chromosome. They are difficult to identify when in solution, but once you trap and stretch them in the focal plane they are beautiful”
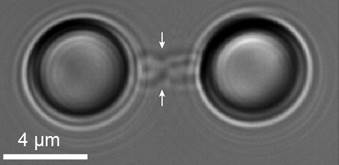
 Diagram of a chromosome bound to microspheres trapped in the laser of an optical tweezer. (right) Microscopy image of a chromosome trapped in an optical tweezer.")
(left) Diagram of a chromosome bound to microspheres trapped in the laser of an optical tweezer. (right) Microscopy image of a chromosome trapped in an optical tweezer.
Once you catch a chromosome, the first and most straightforward experiment one can do is to pull on the chromosome by moving the two beads apart. This will apply a force to the chromosome, which we can measure by detecting how much the beads move out of the center of the optical traps. In these experiments we observed two striking things: the mechanical response varied dramatically from chromosome to chromosome, with some being much stiffer or softer than others and, secondly, the mechanical response was not linear, but instead clearly showed stress stiffening – the more you pull on the chromosome, the harder it gets to pull more. These observations are not particularly unexpected – it is known that different chromosomes vary drastically in size and most biological polymers, including DNA, show non-linear mechanics – but once you look a bit closer, there were many fascinating and unexpected details

So, why do biopolymers get stiffer the more you pull on them? If we consider a single polymer strand held at a fixed end-to-end distance, then this chain can take many different conformations, it will show thermal fluctuations and explore these different conformations. When the ends of the chain are pulled further apart, the number of possible conformations decreases, which means there is an entropic cost against extending the polymer, which leads to a restoring force resisting against stretching. As the chain nears full extension, this force starts to diverge, i.e. the force goes to infinity, making it impossible to ever pull a polymer into a completely straight conformation.
To explain why our observation was so remarkable, we have to do a bit of math (remember the quote by Ian Hickson). Generally speaking, the force F is expected to diverge with the extension d of the polymer according to F ~ (d-dc)-δ , where dc is the maximal extension and the exponent δ is positive. If we now calculate the stiffness K = (dF)/(dd) of such a polymer, we get K ~ (d-dc)-(d+1), which we can combine with the first equation to get K ~ F(d+1)/d. The exponent γ = (δ+1)/δ is called the stiffening exponent. Now, we know that δ > 0, so therefore it is immediately clear that γ always has to be larger than 1. However, when we took our chromosome stretch curves and calculated their slope (the experimental stiffness), we found that on average γ = 0.82±0.05, clearly smaller than 1. Now, as we just explained, this should be impossible. Of course, our first reaction was that we had done something wrong, either in the data analysis or in how we conducted the experiments. So, we double and triple checked our analysis, we performed numerous additional experiments, but after the dust settled we still got the same (impossible) result of γ < 1.
How can we now reconcile theory and experiment? It is impossible for a single polymer strand to have γ < 1, but how about many polymer strands? It turns out, if you have multiple polymer strands in a sequence, which all show stress-stiffening, and if the properties of these polymers are distributed just right, then this collective of polymers can have much weaker stress stiffening than the individual polymers. What is happening, is that these polymers stiffen one after the other: First the response is dominated by the softest polymer, but once it stiffens, the next strand becomes the softest and so on, leading to a combined response with weaker stiffening than each individual polymer. The saying goes ‘a chain is only as strong as it’s weakest link’, but in this case, which link is the weakest is changing as we stretch the chromosome.
But exactly how crazy is it to think of the chromosome as a series of polymers with different properties? Actually, not all that crazy. When chromosomes are stained in a specific way (the so-called Giemsa staining) they show a clear striping pattern, which makes it very plausible that these parts also have different mechanical properties. In this way the heterogeneous structure of the chromosome determines how mitotic chromosomes react when they are stretched.
So, what is coming next for this method? Of course, these observations got our scientific minds cooking: What are these substructures? What do all the proteins (condensins, cohesins, etc.) in the chromosome do? Can we image the chromosomes in interesting ways? A million new research questions worth exploring. While the chromosomes in our experiments are pulled by the chromosome ends (telomeres) with optical tweezers, in cells during mitosis, the mitotic spindle segregate chromosomes by pulling from the center (centromeres). During this process the spindle applies considerable pulling force on chromosomes and one exciting adaption of our system would be to mimic this process and pull chromosomes apart by the centromere using optical tweezers. This would allow detailed single chromosome analysis of sister chromatid segregation and the enzymes involved. One can also imagine evolving our system in order to identify and analyze specific chromosomes. This would enable us to pair the structural characteristics of individual chromosomes with their mechanical properties and get an even better understanding of chromosome biology. It would also make it possible to study chromosome specific syndromes like fragile X syndrome, where the X chromosome has a structural defect. This could drastically improve diagnosis of this syndrome, which is the most common cause of severe intellectual disability.
However, the long game for this method is hopefully not in research labs but in clinical settings. Many forms of cancer are related to chromosomal errors and instability during formation or separation of mitotic chromosomes.This is also the major cause of spontaneous miscarriage during pregnancy. Hopefully, once our method is mature enough, it can escape the research lab to have a broader impact.
Follow the Topic
-
Nature
A weekly international journal publishing the finest peer-reviewed research in all fields of science and technology on the basis of its originality, importance, interdisciplinary interest, timeliness, accessibility, elegance and surprising conclusions.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in