Beyond the target: how a Parkinson's drug doubles up as an inducer of microbial iron starvation
Published in Microbiology and Pharmacy & Pharmacology
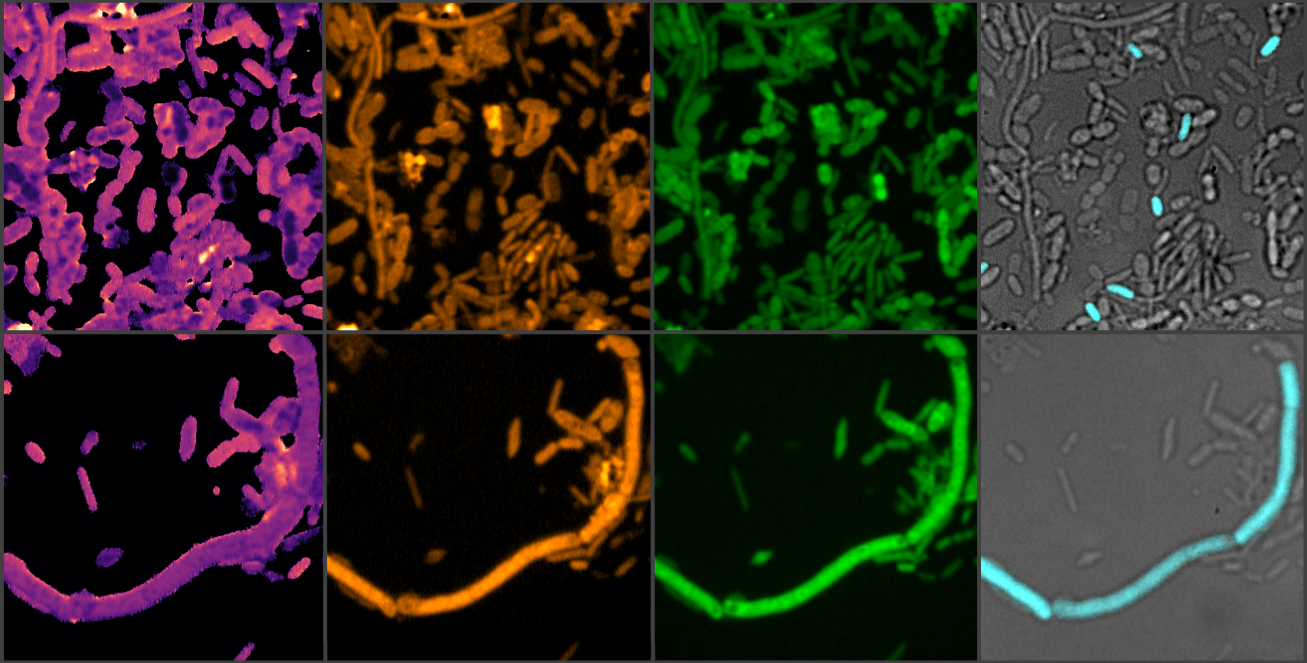
Key to tackling these questions effectively was a highly collaborative team with complementary expertise, led by Michael Wagner (University of Vienna) and Ji-Xin Cheng (Boston University), and myself. Since 2019, our team has been working together on the development of a Raman spectroscopy-based platform to measure microbial activity at the single-cell level with high-throughputs in complex microbial communitiesref2. This platform, termed SRS-FISH, was an ideal tool to get our project off the ground and to study the effects of drugs on whole microbiome communities. This platform will surely be the basis for many more projects to come from this team – fortunately, Michi’s curiosity and questions never seem to end.

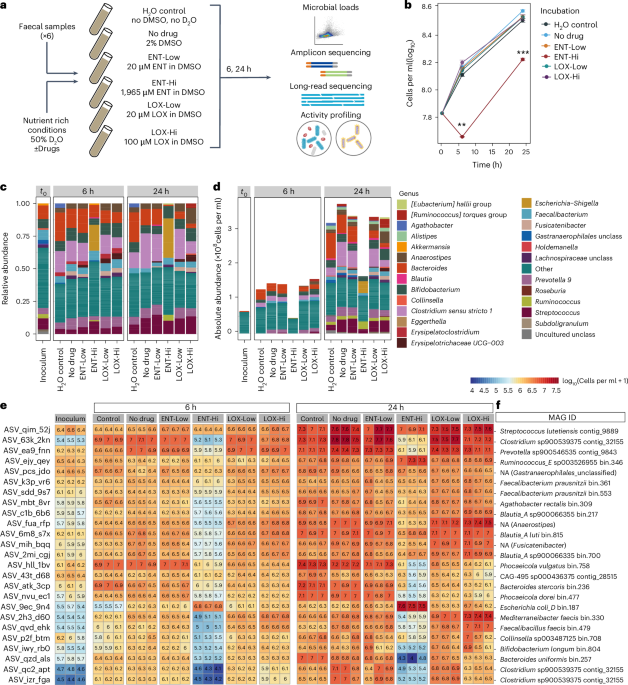
We started our project by recruiting a group of healthy people (mostly colleagues or friends) to provide a faecal sample, a good proxy for the distal gut microbiome and a non-invasive way to access these microbial communities. From the collected samples, we prepared a faecal slurry, distributed it into vials, supplemented each vial with the drug of interest and sampled it over a 24-hour period. Among drugs targeting the nervous system, we chose to focus on entacapone (Parkinson's disease adjuvant drug reducing the breakdown of the mainstay medication levodopa outside the brain) and loxapine succinate (to treat schizophrenia). These drugs were selected because of their established clinical use and documented strong inhibitory effects on microbial growth, with some selectivity for specific gut microbes. By the time we reached the second sampling point, we already suspected that entacapone affected the microbial community: cell pellets from the entacapone-treated samples were visibly smaller than those from the control or loxapine-treated samples. Microbial cell counts and sequencing results confirmed our suspicions: entacapone reduced total microbial loads and had a pronounced effect on microbiome composition. Loxapine also affected the microbiota, but to a much lesser extent. To determine how these drugs impact microbial activity, we supplemented our incubation vials with heavy water (D2O), a tracer for metabolic activityref2. This allowed us to follow microbial activity at the single cell level using SRS-FISH - thank you Jannie and Klara - I remember that at one point we had more FISH labelling experiments running in parallel than timers available in the lab!
After many shipments to Boston - none of which resulted in lost or damaged samples, arguably one of our study’s greatest achievements - we observed changes in the activity of specific microbial community members even when these were supplemented with low drug concentrations. Another surprising finding from our stimulated Raman scattering (SRS) analyses was the detection of entacapone bioaccumulation in microbial cells, made possible by the intrinsic photothermal properties of entacapone, which we were able to image with single-cell resolution thanks to the Boston team's curiosity, skills and state-of-the-art optics instrumentation.

With these new findings about entacapone in mind, we turned to a chemist and collaborator, Thomas Boettcher, to help us track the fate of this drug in our samples using analytical chemistry techniques. Just one slide into the project presentation – the one slide featuring the entacapone chemical structure - Thomas interrupted with questions about iron concentrations and oxidation state in the vials. This was very confusing at first - why was he talking about metals? - but it all made sense when he explained that the presence of a catechol group probably enabled entacapone to chelate iron present in our incubation vials - potentially blocking microbes from accessing this micronutrient. We left the meeting with a new strategy: add an excess of iron to our incubations, more than what entacapone could chelate based on the stoichiometry of the entacapone:iron complex, and see if we could reverse the effect of entacapone. And indeed, we could! At the same initial meeting, Thomas also noted that the entacapone effect could be potentially circumvented by microbes capable of producing siderophores, molecules that bind iron with very high affinity, higher than entacapone. We then moved on to examine the genomes of microbiome members unaffected by entacapone and found out that these indeed possess the genomic ability to produce high-affinity iron-binding molecules. These observations were only possible due to the high degree of completion of the metagenome-assembled genomes retrieved, enabled by long-read sequencing and by a super skilled and efficient in-house sequencing facility (JMF Vienna) and nanopore wizard Rasmus! This study was indeed a big team effort - it was such a pleasure to work together with so many awesome scientists.
You can read about all details of our study in the full paper here.
Microbiologist turned microbial ecologist; interested in all forms of microbe-microbe and host-microbe interactions in the gut.
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in