Breaking the Efficiency Barrier in Dicot Prime Editing: The Journey with Tomato and Arabidopsis
Published in Microbiology, Protocols & Methods, and Cell & Molecular Biology
Early Challenges and Background
Gene targeting (GT) allows precise gene/allele replacement via homologous recombination (HR) that facilitates genetic information exchange between a target site and a homologous donor template. CRISPR-Cas-based plant GT has been a challenge due to the strong preference for the competitive double-stranded break (DSB) repair pathway called canonical nonhomologous end-joining (cNHEJ) over HR (Puchta, 2017). Even with our efforts to improve GT using a bean yellow dwarf virus (BeYDV) geminiviral replicon system (Baltes et al., 2014) and the CRISPR-LbCas12a nuclease, the efficiency remained lower than what is practical for breeding applications (Vu et al., 2020). As a result, we started exploring other methods for performing precision plant genome engineering, such as prime editing (PE), but initially, we faced challenges as the efficiency was very low (Vu et al., 2022). In the same work, we discovered that the low PE efficiency in tomatoes might be partially attributed to the negative impacts of PBS-spacer self-complementarity and the fusion of Cas9 and RT peptides (Vu et al., 2022).
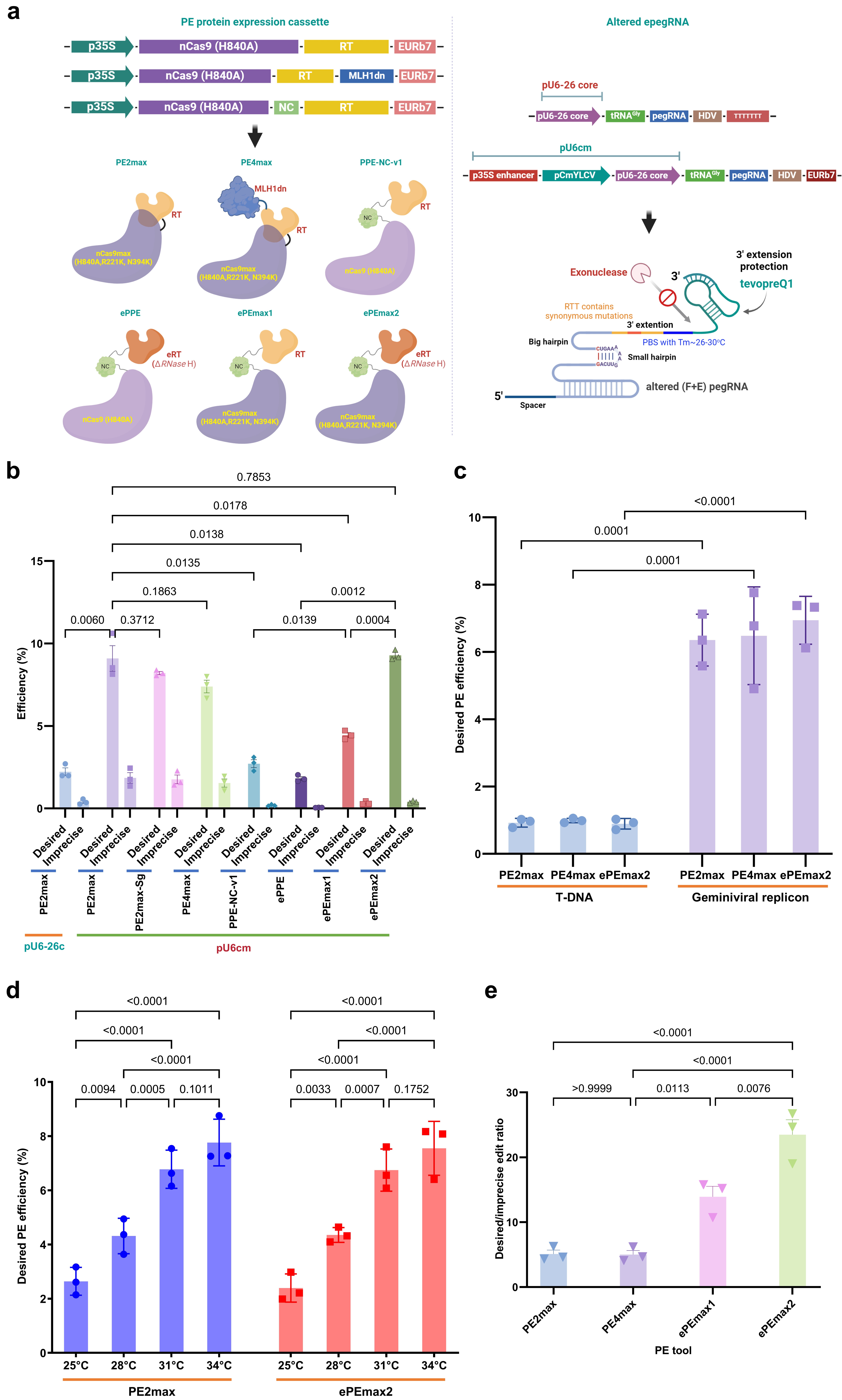
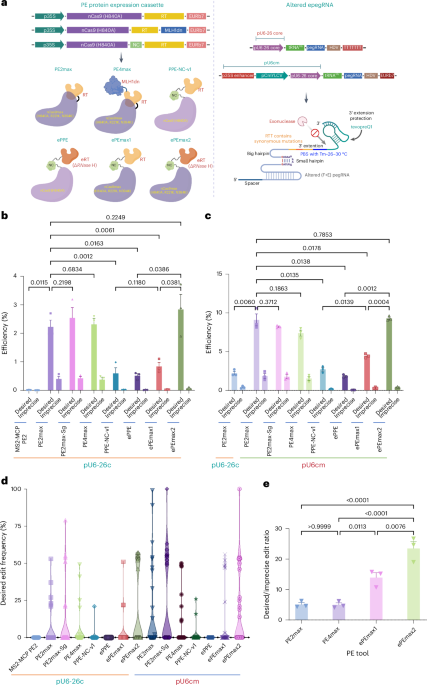
Fig. 1: Recombining the PE tool for breaking the efficiency barriers in tomato. a, The PE protein configurations (left panel) and altered pegRNA variant (right panel). b, Performance of the recombined PE tools using pU6cm for driving the altered pegRNA transcription at the SlALS1 site at the callus stage. c-d, Comparison of T-DNA and the geminiviral replicon in PE efficiency (c) and the impact of heat treatments in PE performance (d) at the SlALS1 site using the PE2max-based tool at the callus stage. e, Desired/imprecise edit ratios. The experiment was conducted in triplicates. The statistical analysis was performed with the GraphPad Prism 9 using a two-sided, uncorrected Fisher’s LSD test, n=3. All the data points are shown on the plots. The error bars represent ± SEM.
Breakthrough in Dicot Prime Editing
In our recent efforts, we engineered synthetic combinations of PE components developed in mammals and monocots to generate novel PE tools (Fig. 1a) that could break the efficiency barrier in tomatoes and Arabidopsis. Our approach involved using an altered epegRNA variant (Nelson et al., 2022), novel combinations of PE protein components (Chen et al., 2013; Chen et al., 2021; Doman et al., 2022; Nelson et al., 2022; Zong et al., 2022), overproduction of pegRNA by a U6 composite promoter (Li et al., 2022) Fig. 1a), employing the geminiviral replicon for amplifying the PE tool and heat treatment (Vu et al., 2020).
Compared to our previous PE system (Vu et al., 2022), we achieved a dramatic improvement in PE efficiency, up to 678-fold, using modified PE tools with U6cm promoter-driven modified epegRNA expression (Fig. 1b). The BeYDV replicon system, known for enhancing mRNA transcription and protein translation, significantly supported our CRISPR-Cas-based PE applications in tomato. We observed up to a 7.9-fold improvement in PE efficiency using the replicon compared to the T-DNA baseline (Fig. 1c). Subsequent analysis by qPCRs and Western blot showed significant increases in the transcription of PE components and PE protein levels delivered by the replicon system, compared to the T-DNA-based tools. Heat treatment further enhanced the PE efficiency (Fig. 1d). The newly engineered ePEmax2 variant showed comparable desired efficiency with PE2max but with a much higher desired/undesired PE efficiency ratio (Fig. 1e). The improved tool was subsequently validated at seven out of ten more loci with eleven pegRNAs (Fig. 2a). The efficiency of the dicot PE tools was comparable or even higher at some sites than that in monocots (Lin et al., 2020; Zong et al., 2022). Further analysis revealed very low off-targeting activities of the engineered PE tools at the tested loci at both the callus and plant stages. The improved PE tools also ensured PE efficiency by paired pegRNAs and multiplexing capability. However, the locus-dependent nature of the engineered PE tool suggests the requirement for further enhancement.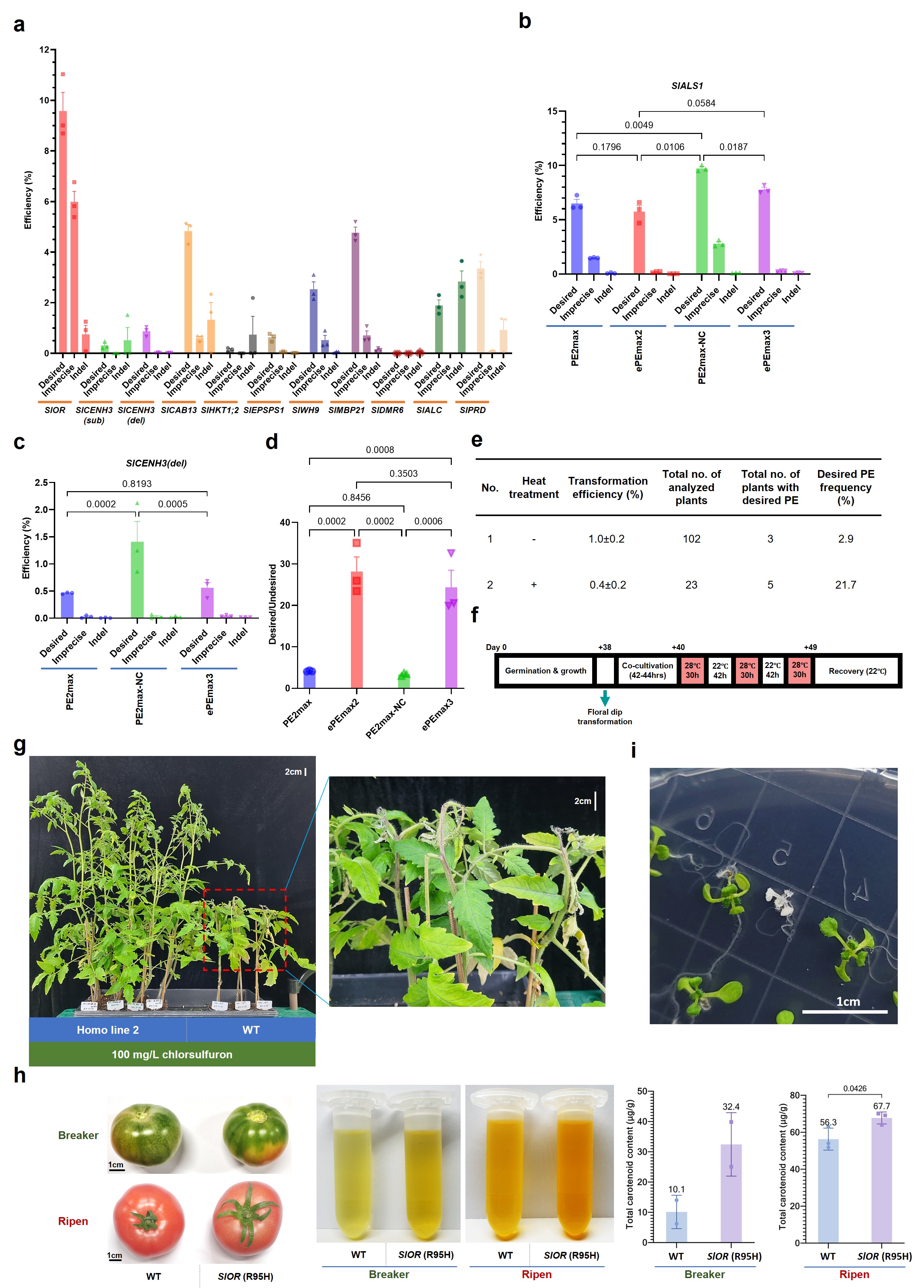
Fig. 2: The enhancement and applications of the engineered PE tools. a, Callus-stage PE efficiency revealed by targeted deep sequencing at other ten tomato loci. b-d, Improving the PE tools using the NC RNA chaperone at the SlALS1 and SlCENH3(del) sites. e-f, Design and employment of the enhanced PE tool in Arabidopsis. e, The PE efficiency obtained with and without heat treatment. f, The heat treatment conditions. g, Herbicide (chlorsulfuron) tolerance of the T1 plants carrying homozygous SlALS1 (P186S) allele. h, The enhancement of total carotenoid production in tomato carrying homozygous SlOR (R95H) allele. i, T2 Arabidopsis seedlings of the AtPDS3 events carrying 19% of the knockout allele (M30stop) germinated on an antibiotic-free medium. The albino plant carried homozygous alleles of AtPDS3 (M30stop). The statistical analysis was performed with the GraphPad Prism 9 using a two-sided, uncorrected Fisher’s LSD test, n=3 (three biological replicates). All the data points are shown on the plots. The error bars represent ± SEM.
Engineering Enhanced PE Tools
The RTT template's secondary structure was considered very important to their competency as the substrate for the RT reaction (Doman et al., 2023). We hypothesized that the RTT secondary structure could contribute to the variation in PE efficiency and be alleviated by an RNA chaperone, such as the nucleocapsid (NC), that enhanced PE efficiency in monocots (Zong et al., 2022). In the initial experiments, the PE2max protein showed the highest activity (sum of precise and imprecise efficiencies). We fused the NC to the C-terminal of PE2max (PE2max-NC variant). We then constructed a similar configuration of the ePPE (Zong et al., 2022) by truncating the RNAse H domain from the RT sequence of PE2max and inserting the NC between the nCas9(H840A) and the PEmaxΔRNaseH forming ePEmax3. Our results showed that PE2max-NC and ePEmax3 could significantly enhance the efficiency and accuracy of PE (Fig. 2b-d). This enhancement was evident in both tomatoes and Arabidopsis, with heat treatment further improving PE performance up to seven-fold (Fig. 2e-f).
Achieving Practical Outcomes
We were focused on creating plants that had been edited with gene-editing technology and didn't contain any added genes in the first generation. It was also important to ensure that these changes would be reliably passed down to future plant generations to help create new plant varieties. We were thrilled that we managed to get transgene-free plants in about 14% of the edited events in the first and next generations, which means we could avoid any worries about integrating foreign DNA and facing potential regulatory issues. Also, the plants had a matching pair of the desired PE alleles, which led to some cool changes in how they look and respond to things like herbicides. For example, they became more tolerant to herbicides or ended up with higher levels of carotenoids in tomatoes (Fig. 2g-h) and albinism in Arabidopsis (Fig. 2i).
Conclusion
Our research significantly advances prime editing tools used in dicot plants, resulting in greater efficiency, accuracy, and transgene-free plants with stably inherited edits. These improvements hold great promise for the future of crop gene editing, opening up possibilities for more precise and efficient modifications in species where conventional editing tools have been less effective. Further studies are needed to validate these tools in other plant species and to explore their potential applications in various agricultural and horticultural settings.
References
Anzalone, A.V., Randolph, P.B., Davis, J.R., Sousa, A.A., Koblan, L.W., Levy, J.M., Chen, P.J., Wilson, C., Newby, G.A., Raguram, A., Liu, D.R., 2019. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 576, 149-157.
Baltes, N.J., Gil-Humanes, J., Cermak, T., Atkins, P.A., Voytas, D.F., 2014. DNA replicons for plant genome engineering. Plant Cell 26, 151-163.
Chen, B., Gilbert, Luke A., Cimini, Beth A., Schnitzbauer, J., Zhang, W., Li, G.-W., Park, J., Blackburn, Elizabeth H., Weissman, Jonathan S., Qi, Lei S., Huang, B., 2013. Dynamic Imaging of Genomic Loci in Living Human Cells by an Optimized CRISPR/Cas System. Cell 155, 1479-1491.
Chen, P.J., Hussmann, J.A., Yan, J., Knipping, F., Ravisankar, P., Chen, P.F., Chen, C., Nelson, J.W., Newby, G.A., Sahin, M., Osborn, M.J., Weissman, J.S., Adamson, B., Liu, D.R., 2021. Enhanced prime editing systems by manipulating cellular determinants of editing outcomes. Cell 184, 5635-5652 e5629.
Doman, J.L., Pandey, S., Neugebauer, M.E., An, M., Davis, J.R., Randolph, P.B., McElroy, A., Gao, X.D., Raguram, A., Richter, M.F., Everette, K.A., Banskota, S., Tian, K., Tao, Y.A., Tolar, J., Osborn, M.J., Liu, D.R., 2023. Phage-assisted evolution and protein engineering yield compact, efficient prime editors. Cell 186, 3983-4002 e3926.
Doman, J.L., Sousa, A.A., Randolph, P.B., Chen, P.J., Liu, D.R., 2022. Designing and executing prime editing experiments in mammalian cells. Nat Protoc 17, 2431-2468.
Li, J., Chen, L., Liang, J., Xu, R., Jiang, Y., Li, Y., Ding, J., Li, M., Qin, R., Wei, P., 2022. Development of a highly efficient prime editor 2 system in plants. Genome Biology 23, 161.
Lin, Q., Zong, Y., Xue, C., Wang, S., Jin, S., Zhu, Z., Wang, Y., Anzalone, A.V., Raguram, A., Doman, J.L., Liu, D.R., Gao, C., 2020. Prime genome editing in rice and wheat. Nat Biotechnol 38, 582-585.
Nelson, J.W., Randolph, P.B., Shen, S.P., Everette, K.A., Chen, P.J., Anzalone, A.V., An, M., Newby, G.A., Chen, J.C., Hsu, A., Liu, D.R., 2022. Engineered pegRNAs improve prime editing efficiency. Nat Biotechnol 40, 402-410.
Puchta, H., 2017. Applying CRISPR/Cas for genome engineering in plants: the best is yet to come. Current Opinion in Plant Biology 36, 1-8.
Vu, T.V., Nguyen, N.T., Kim, J., Das, S., Lee, J., Kim, J.-Y., 2022. The obstacles and potential solution clues of prime editing applications in tomato. BioDesign Res 2022, 0001.
Vu, T.V., Nguyen, N.T., Kim, J., Hong, J.C., Kim, J.-Y., 2024. Prime editing: Mechanism insight and recent applications in plants. Plant Biotechnology Journal 22, 19-36.
Vu, T.V., Roy Choudhury, N., Mukherjee, S.K., 2013. Transgenic tomato plants expressing artificial microRNAs for silencing the pre-coat and coat proteins of a begomovirus, Tomato leaf curl New Delhi virus, show tolerance to virus infection. Virus Research 172, 35-45.
Vu, T.V., Sivankalyani, V., Kim, E.-J., Doan, D.T.H., Tran, M.T., Kim, J., Sung, Y.W., Park, M., Kang, Y.J., Kim, J.-Y., 2020. Highly efficient homology-directed repair using CRISPR/Cpf1-geminiviral replicon in tomato. Plant Biotechnol J 18, 2133-2143.
Zong, Y., Liu, Y., Xue, C., Li, B., Li, X., Wang, Y., Li, J., Liu, G., Huang, X., Cao, X., Gao, C., 2022. An engineered prime editor with enhanced editing efficiency in plants. Nat Biotechnol 40, 1394-1402.
Follow the Topic
-
Nature Plants
An online-only, monthly journal publishing the best research on plants — from their evolution, development, metabolism and environmental interactions to their societal significance.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in