Chromatin Engineering as a Tool to Enhance Bone Regeneration
Published in Bioengineering & Biotechnology


Stem cell plasticity which is fostered by the 3D chromatin conformation, and epigenetic modifications, among other molecular signatures, makes the process of differentiation highly stochastic without external cues. As a result of this intrinsic plasticity of stem cells, direct differentiation towards the cell type of interest becomes particularly challenging. Interestingly, engineered micro- and nano-topographies can result in contact guidance-induced nuclear deformation, and can potentially modulate cellular plasticity and maximize lineage-specific differentiation. However, engineering topographies to regulate phenotypic outcomes for drug/cytokine-free acceleration of tissue regeneration, particularly at the tissue-implant interface has not been investigated.
Chromatin packing dictates the conformation of the 2-meter-long DNA polymer within a relatively small ~10 μm cell nucleus and is a key regulator of transcriptional responsiveness to external cues. Consequently, nuclear morphology plays a critical role in regulating chromatin remodeling and transcriptional reprogramming that have significant implications for the phenotypic outcomes of cells. In vivo, various biological processes such as malignant cell migration and smooth muscle cell contraction, as well as stem cell homing are accompanied by nuclear deformation. Such alterations in nuclear morphology can be replicated on surfaces with defined topographies via contact guidance in vitro. Herein, we integrated micro-topography engineering, molecular biology, and advanced nanostructural imaging tools to investigate the influence of nuclear morphology on the osteogenic differentiation of human mesenchymal stromal cells (hMSCs). Our work demonstrates chromatin engineering via contact guidance-induced nuclear constriction as a tool to modulate cell plasticity and transcriptional responsiveness of stem cells to external differentiation cues. We provide direct evidence for contact guidance-induced chromatin reprogramming and targeted cell differentiation both in-vitro and in-vivo. We show that the resulting chromatin reorganization causes transcriptional reprogramming Furthermore, we demonstrate that microtopography engineering can be utilized as a tool to decouple the effects of the nucleus and cytoskeleton on 3-D chromatin conformation in deformed nuclei. Importantly, we demonstrate, for the first time, enhanced bone regeneration in-vivo due to contact guidance-induced chromatin reprogramming on micropillars.
How nuclear morphology change affect nuclear structure?
To control nuclear morphology, we designed and fabricated micropillar structures with various sizes, shapes, and spacing with methacrylated poly (octamethylene citrate) (mPOC), a citrate-based biomaterial (CBB) with one formulation, CitregenTM, has been used for the manufacturing of biodegradable tissue fixation devices cleared by the U.S.A. Food and Drug Administration (FDA) for use in musculoskeletal surgeries. The microstructures affected cell morphology and cytoskeleton assembly. The F-actin and intermediate filaments provided compressive and pulling force that insert the nucleus in-between the micropillars which resulted in nuclear deformation. Due to the confinement of micropillars, the deformed nucleus showed distinct assembly of nuclear lamin and chromatin density (Fig. 1). Multimerization of lamin a/c happened around the micropillars in deformed nucleus, but at basal side in undeformed one. Consequently, the organization of lamin-associated chromatin is altered in micropillars. We found that the chromatin concentration, especially the heterochromatin-like domains, is increased in hMSCs nuclei on micropillar surfaces when compared to flat surfaces which suggests an increase in their relocalization toward nuclear edge.
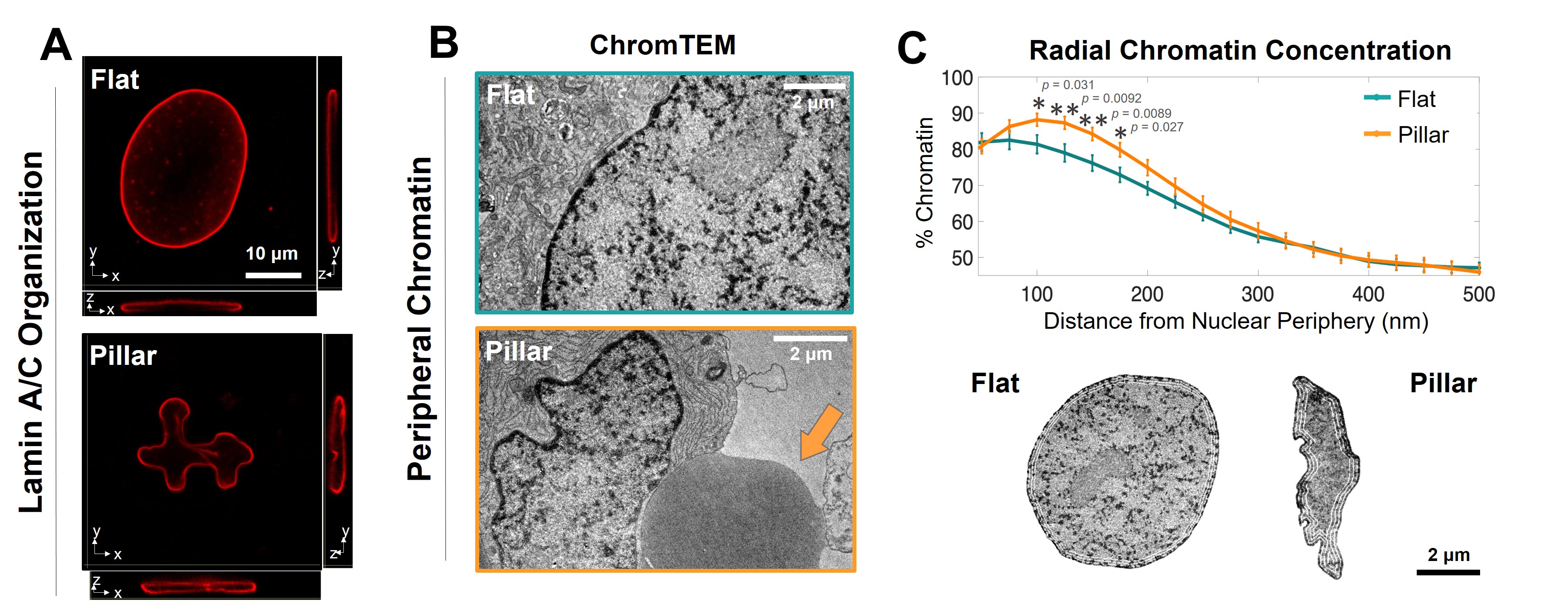
Figure 1. Micropillar-induced nuclear deformation is associated with alterations in nuclear structural components. A. Lamin A/C staining images of hMSC nuclei on flat and micropillar surfaces. B. ChromTEM images of 50 nm thick resin sections of hMSCs seeded on flat and micropillar surfaces. The orange arrow shows the location of micropillars. C. Top: Radial Chromatin concentration analysis defined by the percentage of chromatin within consecutive 25 nm thick bands as a function of the distance from its location to the nuclear periphery. Bottom: ChromTEM images show radial bands at 25 nm, 250 nm, 500 nm, and 750 nm.
How chromatin reprogramming affect transcription?
Based on our previous study1, we reveled that chromatin packing scaling (D) is a key physical property that is associated with transcription regulation and is also a crucial regulator of phenotypic plasticity. Using partial-wave spectroscopic (PWS) microscopy, we are able to measuring D with sensitivity to length scales as small as 20 nm in living hMSCs (Fig. 2). It showed a dramatic decrease of D on micropillars compared to flat surfaces indicating a drastic change in chromatin conformation. To understand how chromatin conformation affect cell phenotype, we performed RNA-sequencing and ATAC-sequencing in hMSCs that were cultured on flat and micropillar surfaces with growth and osteogenic induction medium, respectively. It revealed that the changes in gene expression on micropillars are associated with alterations in chromatin accessibility. Specifically, we found upregulation in the expression of genes involved in response to external stimulus. Thus, we evaluated the lineage-specific responsiveness coefficient, RLS, defined as the average transcriptional response to an external differentiation stimulus of stem cells on a micropillar surface compared to a flat surface. Based on our chromatin packing macromolecular crowding (CPMC) model1, we reveled that the lineage-specific transcriptional response to osteogenic differentiation induction increased in low D cells on micropillars compared to high D cells cultured on flat surfaces indicating enhanced transcriptional compliance. It was further confirmed with in vitro osteogenic differentiation of hMSCs. Cells with deformed nuclei showed enhanced osteogenic differentiation upon induction.
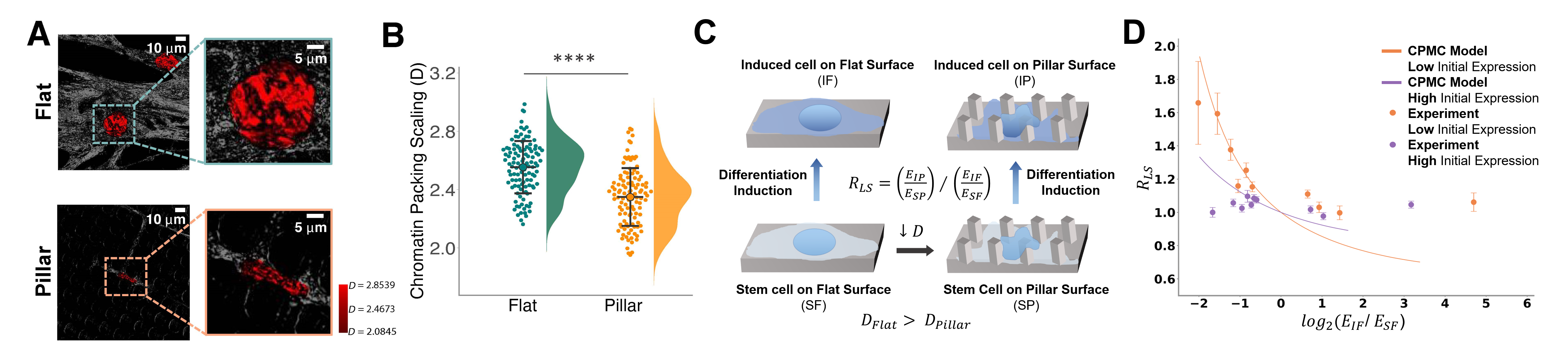
Figure 2. A decrease in chromatin packing scaling in deformed nuclei enhances the responsiveness of hMSCs to osteogenic differentiation. A. PWS microscopy of hMSCs on flat and micropillar surfaces. B. Micropillars deform cell nuclei and significantly decrease average D in stem cells. C. hMSCs seeded on a flat surface (high D) and micropillar surface (low D) (SF and SP, respectively) were induced to differentiate towards osteogenic lineage (IF and IP, respectively). D. Lineage-specific transcriptional response due to differentiation induction increases in low D cells.
Biomedical implication of micropillar implants
To investigate the clinical relevance of our approach, we implanted hMSCs seeded mPOC scaffolds in nude mice with critical-sized calvarial defects (Fig. 3). μCT scans showed enhanced new bone formation treated with micropillar implants. Histology images confirmed the better regeneration efficacy of micropillar implants. Furthermore, a lower D value was observed with micropillars implanted tissue sections indicating the nuclear deformation and chromatin conformation alteration persists in vivo.
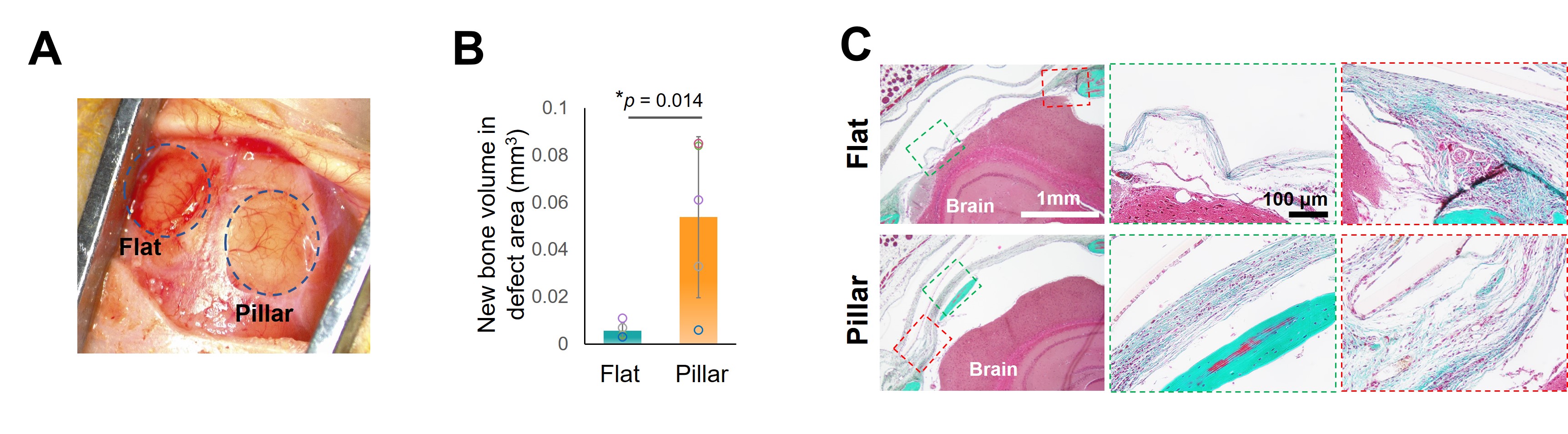
Figure 3. Micropillar-induced nuclear deformation promotes bone regeneration in vivo. A. Two critical-sized cranial defects with a diameter of 4 mm were created, followed by implantation of hMSC-seeded flat and micropillar mPOC scaffolds. B. Newly formed bone volume in the defect region treated with flat and micropillar implants. C. Trichrome staining of calvarial defect tissue treated with flat and micropillar implants.
Overall, surface topology-induced nuclear constriction may serve as a tool for chromatin engineering to predictably modulate the regenerative potential of stem cells. More importantly, contact guidance-induced nuclear constriction promoted bone regeneration in vivo, without using exogenous signaling molecules, highlighting the applicability of microtopography engineering to biomedical implants and devices. Our future step is to develop large scale micro-engineered implants for clinical translation.
References
1. Virk, R. K. A. et al. Disordered chromatin packing regulates phenotypic plasticity. Sci. Adv. 6, eaax6232 (2020).
Follow the Topic
-
Nature Biomedical Engineering
This journal aspires to become the most prominent publishing venue in biomedical engineering by bringing together the most important advances in the discipline, enhancing their visibility, and providing overviews of the state of the art in each field.

Related Collections
With Collections, you can get published faster and increase your visibility.
Implantable wireless communication technologies
Publishing Model: Hybrid
Deadline: Nov 28, 2026
Medical Ultrasound: Emerging Techniques and Applications
Publishing Model: Hybrid
Deadline: Jan 29, 2027



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in