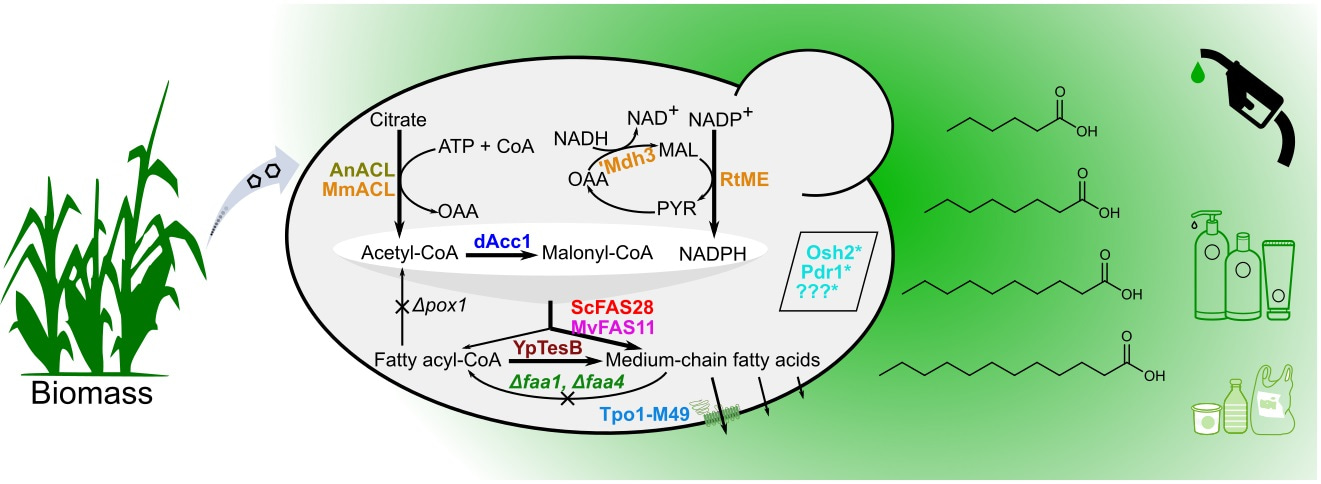
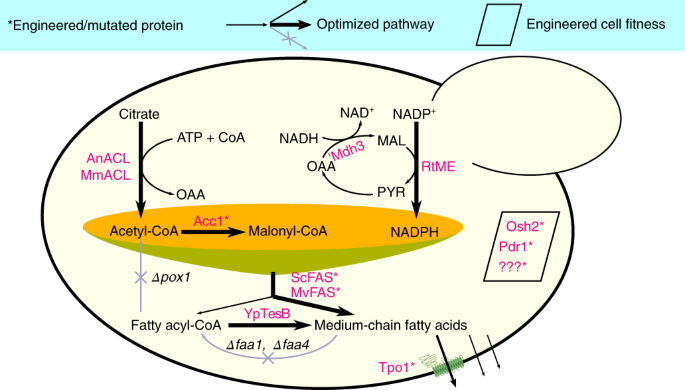
Medium-chain fatty acids (MCFAs, C6–12) and their derivatives are highly desirable chemicals used in many scenarios, such as the production of cosmetics, detergents, plasticizers, lubricants and also biofuels. Instead of petroleum-based synthesis, the coconut and palm kernel oils containing a high fraction of MCFAs (mainly C12) are the natural sources of these fatty acids. The demands on these fatty acids are constantly increasing, and as a significant increase in the plantation of oil plants will re-direct land use for food production there is interest in novel sustainable solutions.
We therefore recruited the microorganism Saccharomyces cerevisiae, generally referred to as Baker’s yeast that has been used by human beings for thousands of years, for production of MCFAs. The native yeast cells only synthesize long-chain fatty acids (C16–18), because the key enzymes, fatty acid synthases, preferentially join 8–9 two-carbon units which are derived from the breakdown of sugars to form long-chain fatty acids. Our previous efforts on embedding short-chain thioesterases into reaction chambers of fatty acid synthase complexes showed that it is feasible to include foreign thioesterase activity into the fatty acid synthesis reaction scheme to hydrolyze shorter chain acyl-ACP intermediates. However, this strategy only enables the engineered yeast producing a small amount of MCFAs[1], so other beneficial mutations[2] were further incorporated into these FAS variants to augment their ability to generate MCFAs. Moreover, engineering the similar FAS enzyme from Mycobacterium to produce MCFAs succeeded as well, and combinatorial expression of both FAS systems synergistically produced more MCFAs.
The second issue we had to overcome was the toxicity of these MCFAs. Although their toxicity mechanism is not clear, these lipophilic compounds are able to readily diffuse across the membrane and induce acidification of the cytosol, or incorporate into membranes and result in membrane damage and leaking. The membrane transporter Tpo1 is involved in cellular resistance against C10 fatty acids. We therefore used a directed evolution strategy, a protein engineering approach where a large number of mutants were screened to get better-performing variants. Based on this we identified a mutant containing changes in three amino acid residues that was able to confer better cellular growth under C10 fatty acid stress as well as promote higher MCFA production. We subsequently used a parallel strategy, adaptive laboratory evolution, to obtain tolerant strains. We chose a starting strain deficient in fatty acid-degrading β-oxidation, which is highly sensitive to the presence of MCFAs during the adaptive laboratory evolution process. After letting the starting strain growing in a medium with C8 fatty acid for more than 100 generations, we obtained a few isolates that had increased tolerance towards C8 fatty acid and were also demonstrated to produce more MCFAs. We then incorporated the engineered fatty acid synthase enzymes and the membrane transporter Tpo1 M49 variant, and other metabolic manipulation into one of the tolerant strains to enable efficient production of more than 1 g/L MCFAs from 20 g/L of glucose, and a 250-fold improvement over the original yeast strain (See Figure 1 for more details).

Figure 1. Multidimensional metabolic engineering of yeast cell factory for efficient synthesis of medium-chain fatty acids.
In specific chassis cells used for the production of chemicals, metabolic engineering implements a range of genetic modifications, most of which are relevant to cellular enzymes and therefore intended to augment the metabolic flux towards desired products. Although the advances in molecular biology and synthetic biology have already enabled dramatically rapid construction of biosynthetic pathways for the products of interest, improving the production phenotypes in terms of titer, rate and yield (TRY) of desired products are the most challenging work, but the prerequisite for the commercialization of a specific bioprocess[3]. TRY traits are quantitative or complex traits that are determined by the cumulative actions of many genes and the environment, and it is therefore generally difficult to improve the TRY metrics of an engineered strain. Optimizations at multiple levels or scales, for instance the combinatorial engineering to improve the activities and properties of key enzymes, to tune the expression of enzymes lie in the synthesis route, to rewire the central metabolism to provide essential precursors but not introduce burdens on cellular growth, and to increase the cellular tolerance to toxic substrates or products, are often needed. Our work is a successful example of combining protein engineering, pathway engineering and tolerance engineering to improve the production of chemicals that are toxic to the microbial biocatalyst. Check out our recent paper published in Nature Catalysis[4] to find out how we implement multidimensional metabolic engineering to build yeast cell factories, and also our previous work on medium-chain chemical production[5].
[1] Zhu, Z. et al. Expanding the product portfolio of fungal type I fatty acid synthases. Nat Chem Biol 13, 360-362, doi:10.1038/nchembio.2301 (2017).
[2] Gajewski, J., Pavlovic, R., Fischer, M., Boles, E. & Grininger, M. Engineering fungal de novo fatty acid synthesis for short chain fatty acid production. Nat Commun 8, 14650, doi:10.1038/ncomms14650 (2017).
[3] Nielsen, J. & Keasling, J. D. Engineering cellular metabolism. Cell 164, 1185-1197, doi:10.1016/j.cell.2016.02.004 (2016).
[4] Zhu, Z. et al. Multidimensional engineering of Saccharomyces cerevisiae for efficient synthesis of medium-chain fatty acids. Nat Catal 3, 64-74, doi:10.1038/s41929-019-0409-1 (2020).
[5] Zhu, Z. et al. Enabling the synthesis of medium chain alkanes and 1-alkenes in yeast. Metab Eng 44, 81-88, doi:10.1016/j.ymben.2017.09.007 (2017).
Follow the Topic
-
Nature Catalysis
This journal brings together researchers from across all chemistry and related fields, publishing work on homogeneous catalysis, heterogeneous catalysis, and biocatalysts, incorporating both fundamental and applied studies.


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in