Impaired chromatin remodeling in cranial neural crest cells can evoke orofacial clefts
Published in Biomedical Research

Orofacial clefts are the second-most common birth defect arising in about 1 to 7001. Besides the numerous surgical interventions that are needed, they come along with attendant symptoms such as feeding and breathing impairments, hearing and speech problems, and they often pose a life-long psychological burden on patients. Multiple factors can account for the formation of orofacial clefts such as smoking, alcohol exposure, folate deficiency, zinc deficiency, viral infection and intake of certain medicinal drugs during pregnancy. Besides these environmental factors, orofacial clefts are also caused by genetic factors. They are a feature of many genetic syndromes. Lately, more and more genetic risk factors for non-syndromic orofacial clefts are being identified. Nevertheless, molecular reasons why a distinct genetic variant causes orofacial clefting mostly remain at large.
Most parts of the face emerge from a tissue specific for vertebrates, the cranial neural crest. These fascinating multipotent neural crest cells arise next to the neural tube, proliferate heavily and migrate towards their destinations, mostly the pharyngeal arches. Finally, they differentiate into diverse tissues such as facial bones, cartilage, smooth muscle and cells of the nervous system. Consequently, these cells undergo drastic changes in their transcriptional profiles during their development.
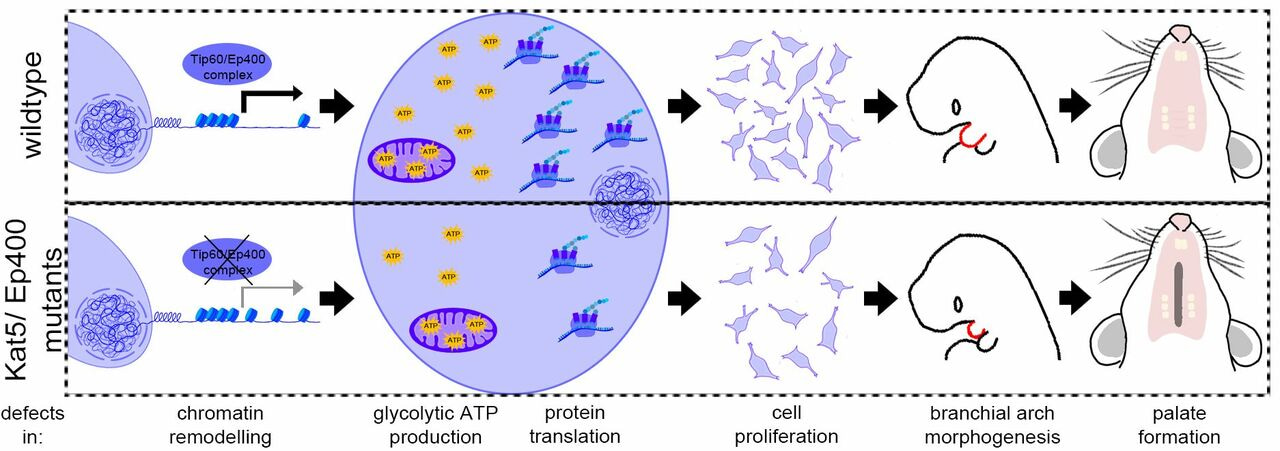
We set out to analyze why mutations in KAT5 (synonym: TIP60) can cause syndromic and non-syndromic orofacial clefts as was identified recently2,3. To this aim, we inactivated the Kat5 gene in a cranial neural crest cell line derived from mouse via CRISPR/Cas9. Kat5 is an acetyl transferase and a component of a multi-subunit chromatin remodeling complex called Tip60/Ep4004. This complex acetylates histones on multiple lysine residues via Kat5. Additionally, with the help of the central ATPase, Ep400, the complex can exchange the canonical histone H2A with the variant H2A.Z in the vicinity of transcriptional start sites. Both processes lead to a more efficient gene expression of target genes.
To get an overview of the genes whose expression is controlled by the Tip60/Ep400 complex in cranial neural crest cells, we also generated Ep400-deficient cells and performed RNA-seq with mutant clones deficient of either gene, Kat5 and Ep400. As expected, deregulated genes were similar in both mutants. Prominent affected cellular processes in both mutants were glycolysis/carbohydrate metabolism, amino acid metabolism and translation, pointing to a role of the complex in maintaining basic cellular functions. Indeed, we found differences in histone acetylation and H2A.Z deposition around transcriptional start sites of genes of each category in our mutant clones. Moreover, glycolytic ATP production and translation rates were actually reduced. It is highly conceivable that these impairments in basic cellular processes lead to the reduced proliferation rates we observed in our mutant clones. To check if the observed effects are also true in the in vivo situation, we created mouse mutants in which Kat5 or Ep400 are deleted specifically in neural crest cells. We detected reduced proliferation and enhanced apoptosis of cranial neural crest cells at early stages such as E9.5 and E10.5, resulting in smaller pharyngeal arches of homozygous mutants. Shortly before birth (E18.5), facial structures were nearly totally absent in homozygous mutants, while heterozygous Kat5 mutants displayed orofacial clefts and a reduced palatal thickness. Thus, even a reduction in protein levels of Tip60/Ep400 complex components can lead to insufficiencies in cranial neural crest cells and finally cause orofacial clefting. This closely resembles the situation in patients where only one allele of KAT5 is mutated. With our model systems, we could relate the occurrence of orofacial clefts to disturbed basic cellular processes in cranial neural crest cells.
Our data show that the two central components Kat5 and Ep400 of the Tip60/Ep400 complex play a prominent role in metabolic reprogramming of cranial neural crest cells and thereby influence survival and proliferation. Interestingly, the activities of Kat5 and Ep400 are themselves regulated by key metabolites such as acetyl-CoA and ATP. Thus, they may sense the metabolic status of cells and relay this information onto gene expression profiles. Therefore, further studies might eventually unveil mechanisms helping to manipulate cranial neural crest cells in a therapeutically relevant manner.
(We acknowledge Matthias Weider and Theresa Schmid for the help in editing this article)
Read the paper: https://www.nature.com/articles/s41368-023-00222-7
- Mossey PA, Little J, Munger RG, Dixon MJ, Shaw WC. Cleft lip and palate. Lancet (London, England). 2009;374(9703):1773-85.
- Humbert J, Salian S, Makrythanasis P, Lemire G, Rousseau J, Ehresmann S, et al. De Novo KAT5 Variants Cause a Syndrome with Recognizable Facial Dysmorphisms, Cerebellar Atrophy, Sleep Disturbance, and Epilepsy. Am J Hum Genet. 2020;107(3):564-74.
- Yang J, Yu X, Zhu G, Wang R, Lou S, Zhu W, Fu C, Liu J, Fan L, Li D, Shao Q, Ma L, Wang L, Wang Z, Pan Y Integrating GWAS and eQTL to predict genes and pathways for non-syndromic cleft lip with or without palate. Oral Diseases. 2021;27(7):1747-1754.
- Hota SK, Bruneau BG. ATP-dependent chromatin remodeling during mammalian development. Development. 2016;143(16):2882-97.
Follow the Topic
-
International Journal of Oral Science
The International Journal of Oral Science seeks to publish all aspects of oral science and interdisciplinary fields, including basic, applied and clinical research.

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in