Life history strategies of soil bacterial communities across global terrestrial biomes
Published in Ecology & Evolution

Every student of ecology and evolution has heard of life history strategies. Personally, I have always been fascinated by this idea.
Life history strategy schemes aim to provide a framework for classifying and comparing organisms based on features that translate their adaptation to specific habitats. The development of such a framework is a long-standing quest in ecology science, which T. R. B. Southwood compared to the elaboration of the periodic table in chemistry (Presidential Address to the British Ecological Society in 1977)1. Since then, understanding the life history strategies of organisms from different groups has stimulated many researches, with some famous works coming from plant ecology. One of these was the development by P. Grime of a framework based on three strategies for plants: Competitor, Stress Tolerant and Ruderal (the CSR triangle), classifying organisms according to their adaptation to gradients of stress, disturbance and competition intensity2. Following this work, ecologists have increasingly embraced the idea that measurable traits should be used to compare the strategies of organisms3. This was the beginning of trait-based approaches in ecology which made it possible to identify some key, measurable, trait dimensions that differentiate overarching plant strategies3,4.
During my PhD, life history schemes of plants and soil microbes highly helped me to frame my research hypotheses5,6. However, I also realised that these theories were far less advanced for soil microbes than for plants. They were still mainly based on syntheses of scattered empirical evidences7–11, with no clear consensus. A characterisation of the dominant trait dimensions, using a global dataset, as it has been done for plant4, was lacking.
Therefore, during my postdoc, I wanted to address this knowledge gap!
Looking at available global datasets, I found the study by Bahram et al. 201812, which seemed to fit with what we needed. Their soil samples covered most of the global environmental range across biomes and they sequenced their metagenomes. The use of the omic approach to study microbial traits was beginning to be used in some studies, including some by my new colleagues at the University of California Irvine13,14. So I jumped into the adventure.
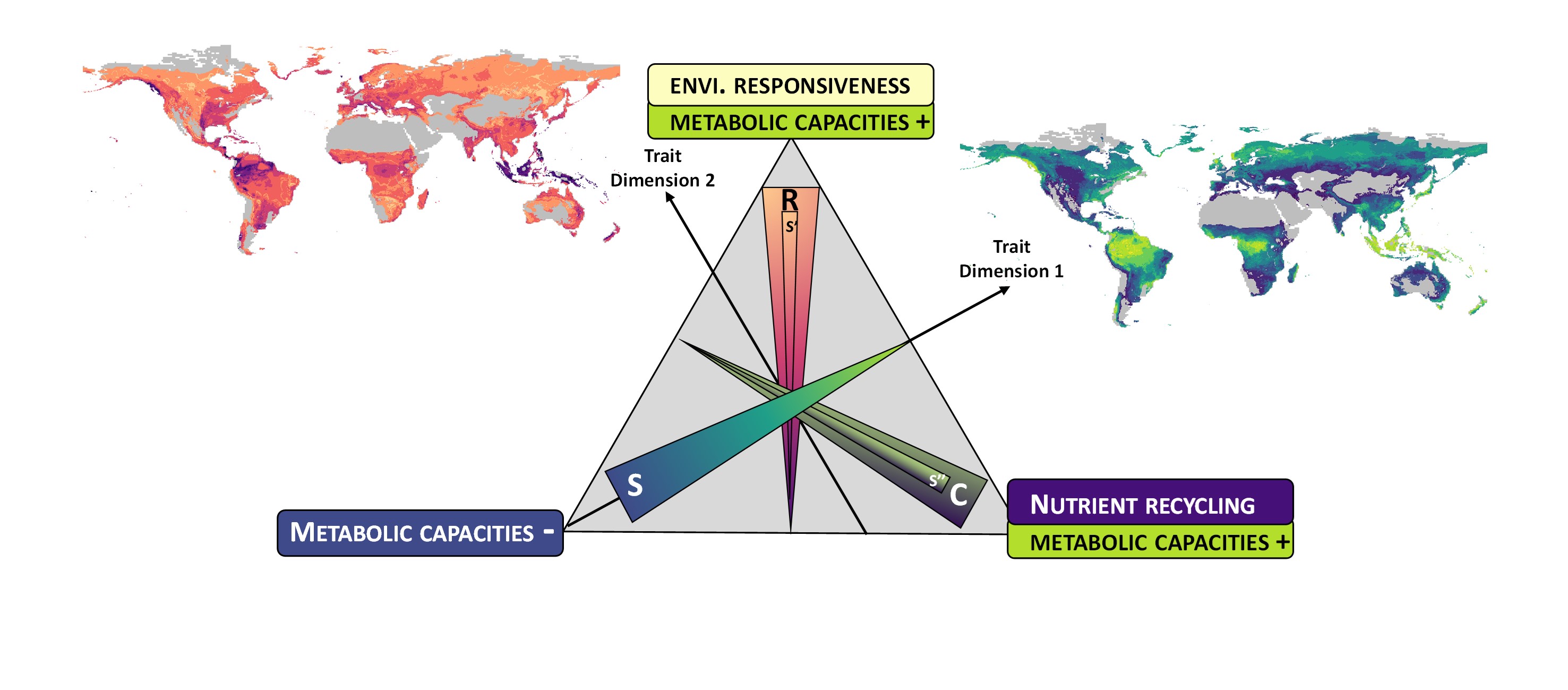
In this study now published in Nature Microbiology, we decided to focus on bacteria that dominate the soil metagenomes. Our first goal was to define and calculate metagenomic metrics that we expected to capture the trait dimensions associated with the life history strategies of soil bacteria. We compiled this list of traits, based on previous syntheses7,10,11 and other studies relating these traits to specific genomic features (see Extended Data Table 1). The idea was also to provide a list of traits that could be used in future studies.
Second, we used an ordination method to describe the main trait dimensions that capture the dominant variation in bacteria life history strategies. It was very exciting to see that the patterns emerging from this analysis showed a triangle (Figure below), as stated by the CSR theory of life history strategies2. The first dimension that shapes this triangle showed a range from bacterial communities with streamlined genomes and simple metabolisms to larger genomes and expanded metabolic capacities, as reported across bacteria isolates15. As metabolic capacities expanded, we observed that bacterial communities became increasingly differentiated along a second dimension, which seems to reflect a trade-off between increasing capacities for environmental responsiveness or efficient nutrient recycling. Finally, we assessed how the position of bacterial communities along these trait dimensions was related to global environmental gradients. Our results showed that soil pH, C:N as well as precipitation level and seasonality were key drivers of bacterial life history strategy, consistent with the theory that resource acquisition constraints, as well as stress intensity and variability play a key role in shaping life history strategy11,16. Bacteria with small genomes were favoured in neutral soils exposed to high water stress intensity and seasonality confirming recent evidence that genome and metabolic streamlining may be a central aspect of the stress-tolerant strategy of soil bacteria17,18. Under more acidic conditions, combined with water stress (low precipitation level and high seasonality), bacteria showed an expanded metabolism oriented towards environmental responsiveness, a profile that we associated with the ruderal strategy. At the third end of the triangle, in acid soils combined with wet and stable precipitation, bacteria showed an expanded metabolism oriented towards nutrient recycling capacities, a profile fitting with competitor strategy.

Overall, our studies have used metagenomes to describe the dominant trait dimensions associated with life history strategies of soil bacteria. This provides a new framework for studying and comparing soil bacterial communities, but does not close the case on soil bacterial life history strategies. Many fascinating questions remain open: have we captured the full spectrum with our community-aggregated traits? Are genomic traits also highly variable within each community? To what extent are genomic traits associated with realised phenotypic traits and performance of soil bacteria at the community and isolate level? Is it possible to transfer our global trait dimensions to local gradients and temporal patterns?...
To conclude this story behind the paper, I would like to share some personal feedback. Although the idea that we developed into this study was clear from the beginning and the original data were already available, it was a long process -about 2 years- to end up with these results. The main challenge was to find the right approach, the right methodology, to keep the exploratory aspect of the study while anchoring it in the theory of trait dimensions and life history strategies. While I was getting my first results, the publication of the paper “Trait dimensions in bacteria and archaea compared to vascular plants” by Mark Westoby et al. was a key step that helped us to consolidate our results and to enrich our discussion. The feedback from the reviewers, especially the ones from Kate Buckeridge, has also been very encouraging for us in pushing further the comparison between our results and the CSR life history strategies.
Overall, in the current age of Big Data, conducting this study has really convinced me that our scientific community has a lot to gain from grounding its large-scale analyses in theory.
References
- Southwood, T. R. Habitat, the templet for ecological strategies? Journal of animal ecology 46, 337–365 (1977).
- Grime, J. P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. The American Naturalist 111, 1169–1194 (1977).
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and soil 199, 213–227 (1998).
- Diaz, S. et al. The global spectrum of plant form and function. Nature 529, 167 (2016).
- Piton, G. et al. Using proxies of microbial community-weighted means traits to explain the cascading effect of management intensity, soil and plant traits on ecosystem resilience in mountain grasslands. Journal of Ecology 108, 876–893 (2020).
- Piton, G. et al. Resistance–recovery trade-off of soil microbial communities under altered rain regimes: An experimental test across European agroecosystems. Journal of Applied Ecology (2020).
- Fierer, N. Embracing the unknown: disentangling the complexities of the soil microbiome. Nature Reviews Microbiology 15, 579–590 (2017).
- Fierer, N., Bradford, M. A. & Jackson, R. B. Toward an ecological classification of soil bacteria. Ecology 88, 1354–1364 (2007).
- Ho, A., Di Lonardo, D. P. & Bodelier, P. L. Revisiting life strategy concepts in environmental microbial ecology. FEMS microbiology ecology 93, fix006 (2017).
- Krause, S. et al. Trait-based approaches for understanding microbial biodiversity and ecosystem functioning. Frontiers in Microbiology 5, (2014).
- Malik, A. A. et al. Defining trait-based microbial strategies with consequences for soil carbon cycling under climate change. The ISME Journal 14, 1–9 (2019).
- Bahram, M. et al. Structure and function of the global topsoil microbiome. Nature 560, 233–237 (2018).
- Malik, A. A. et al. Drought and plant litter chemistry alter microbial gene expression and metabolite production. The ISME Journal 1–12 (2020).
- Ustick, L. J. et al. Metagenomic analysis reveals global-scale patterns of ocean nutrient limitation. Science 372, 287–291 (2021).
- Westoby, M. et al. Trait dimensions in bacteria and archaea compared to vascular plants. Ecology Letters (2021).
- Wood, J. et al. Rethinking CSR theory to incorporate microbial metabolic diversity and foraging traits. The ISME Journal 1–5 (2023).
- Liu, H. et al. Warmer and drier ecosystems select for smaller bacterial genomes in global soils. iMeta e70 (2023).
- Simonsen, A. K. Environmental stress leads to genome streamlining in a widely distributed species of soil bacteria. The ISME Journal 16, 423–434 (2021).
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in