Plant tricks symbiotic bacteria for their own good
Published in Microbiology
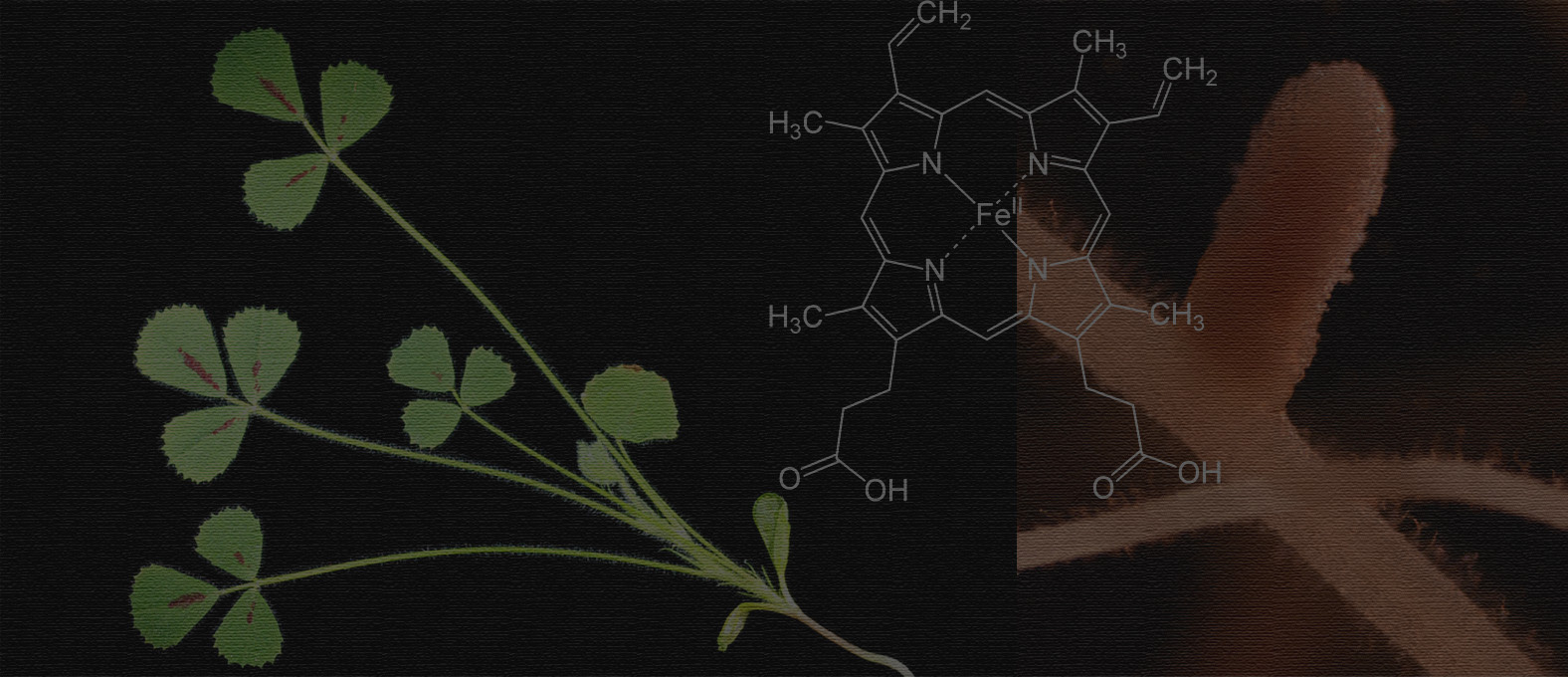
Plants cannot use atmospheric nitrogen and hence depend on soil for fixed nitrogen. The ecologically and agriculturally important symbiosis between rhizobial bacteria and their legume hosts permits these plants to grow without nitrogen fertilizer because the rhizobia use the enzyme nitrogenase to convert atmospheric nitrogen into ammonia. As part of this process, the plant builds specialized guest houses for the bacteria called root nodules and the bacteria are endocytosed into membrane compartments in the cytoplasm of plant cells within the nodules.
In Graham Walker’s lab, I work on understanding the symbiosis between the legume Medicago truncatula (plant) and Sinorhizobium meliloti (bacteria). Interestingly, after entering the plant cell the bacteria undergo a dramatic shift in life cycle called the terminal differentiation process. Bacteria become elongated, and their DNA replicates, but they stop dividing resulting in a long, branched differentiated cell with multiple copies of the genome. They also undergo numerous transcriptional changes; their membrane becomes more permeable and they start fixing nitrogen for the plant. A bacterium that has differentiated cannot return to free-living life even if they are let free into the environment, where they once thrived. What an ordeal these microbes have to undergo and what a cool system to study. A groundbreaking discovery in this system was made in 2010 when Eva Kondorosi, Peter Mergaert and colleagues discovered that this special differentiation process is orchestrated by a set of peptides called Nodule Specific Cysteine Rich peptides1. There are about 700 of these peptides encoded in the M. truncatula genome and the detailed molecular mechanism of action remains to be uncovered.
The Walker lab started working on the shortest cationic NCR peptide-NCR247 (24-amino acids). It was known that the peptide is anti-microbial when used at higher concentrations on various bacteria. However, when used at a more physiological, sublethal concentration, microarray analysis revealed 15% of genes were altered in the transcriptome of S. meliloti many of these changes were comparable to what happens during symbiosis2. I joined the lab as a postdoc and got curious about a set of genes that were regulated by an iron-responsive transcriptional regulator RirA. This drew my attention because of my experience and fascination with understanding iron regulation in host-microbe interaction. I jumped right into it. Bacteria have tightly controlled iron homeostatic mechanisms and strictly regulate their iron intake. Interestingly, I found that treatment with NCR247 increases iron import into the bacteria, even when the bacteria already have sufficient iron for survival. How can a plant peptide drive iron uptake in bacteria? This remained a puzzle for almost a year and I couldn’t find a definitive answer.
Right eyes at the right time: Serendipitously, when working on a completely different aspect of the project I noticed something that solved the peptide iron puzzle and changed the course of my postdoc research. When we overexpressed MBP-tagged NCR247 in E. coli, the protein purified in a reddish-brown color. The presence of reddish color in a protein most likely means there is an iron or iron cofactor associated with it. Through multiple bioanalytical and biochemical assays, I confirmed the presence of an important iron-containing cofactor “heme” in the purified protein. And thus began my adventure in post-doc.
 and Siva Sankari (right)")
I showed that the synthesized NCR247 peptide can bind intracellular heme of S. meliloti. We have extensively characterized the biochemical properties of heme binding to NCR247. How does NCR247 binding heme ultimately result in the import of iron? Iron regulation in S. meliloti is governed by two regulators Irr and RirA3. Interestingly, Irr is regulated by heme3. Through a complex series of events validated in the manuscript, we showed that NCR247 overrides the normal iron homeostatic mechanism in order to trigger the bacteria to import more iron.
Why do bacteria need more iron during symbiosis? The key Nitrogen-fixing enzyme, nitrogenase requires 24-32 units of iron for its structure and functioning and there is excess demand for iron during symbiosis4. NCR peptides are expressed differentially in various zones of nodules and NCR247 is expressed in a zone just ahead of the nitrogen-fixing zone. Since having this excess iron all the time is delirious to the bacteria it makes sense to import iron only when it is needed and when protected in a microaerobic nodule environment where iron is less toxic. It is like the plant saying “Hey bacteria, now is the time to import the excess iron to fix nitrogen for me”. Most importantly, we showed NCR247 is needed for an effective nitrogen-fixing symbiosis.
Over the past decade, free heme has been implicated in the pathology of a breathtakingly diverse variety of genetic and non-genetic human diseases and conditions, including hemolytic genetic disorders such as sickle cell disease and β-thalassemia; diseases caused by infectious agents that result in the release of free heme (e.g., malaria and sepsis); stored blood used to treat trauma-hemorrhage; cancer; kidney disease; immune-mediated inflammatory diseases; cardiovascular disease; ischemia/reperfusion injury; atherosclerosis, neurodegeneration, and Alzheimer’s Disease. Thus, heme has begun to attract attention as a target for therapeutic interventions5. Also, a striking number of important bacterial pathogens6 and eukaryotic parasites7 and worms- some of which infect significant fractions of the world’s population but lack an effective treatment- critically need to import heme from their environment to live. In principle, these pathogens could be controlled if there was a way to limit the free heme in their extracellular environments.
From curious observation to clinical application: Leveraging the unique heme-binding property of NCR247, we picked a few of these issues to test and potentially develop clinical applications. We were successful in developing preliminary evidence for many potential therapeutic applications including inhibiting the growth of some heme-requiring pathogens and parasites, preventing the toxic peroxidase activity of the Aβ-heme complex associated with Alzheimer’s disease, and removing toxic free heme accumulated in stored blood. We are continuing this journey with more exciting discoveries and clinical development. This study is a perfect example of what basic research can bring to the table of the clinical world and throws light on the importance of emphasis on basic research.
- Van De Velde, W. et al. Plant peptides govern terminal differentiation of bacteria in symbiosis. Science (80-. ). 327, 1122–1126 (2010).
- Penterman, J. et al. Host plant peptides elicit a transcriptional response to control the Sinorhizobium meliloti cell cycle during symbiosis. Proc. Natl. Acad. Sci. U. S. A. 111, 3561–3566 (2014).
- O’Brian, M. R. Perception and Homeostatic Control of Iron in the Rhizobia and Related Bacteria. Annu. Rev. Microbiol. 69, 229–245 (2015).
- Brear, E. M., Day, D. A. & Smith, P. M. C. Iron: An essential micronutrient for the legume-rhizobium symbiosis. Front. Plant Sci. 4, (2013).
- Immenschuh, S., Vijayan, V., Janciauskiene, S. & Gueler, F. Heme as a Target for Therapeutic Interventions. Front. Pharmacol. 8, (2017).
- Richard, K. L., Kelley, B. R. & Johnson, J. G. Heme uptake and utilization by gram-negative bacterial pathogens. Front. Cell. Infect. Microbiol. 9, 81 (2019).
- Kořený, L., Oborník, M. & Lukeš, J. Make It, Take It, or Leave It: Heme Metabolism of Parasites. PLoS Pathog. 9, (2013).
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in