Ripping Pathogens to Shreds: How a Cellular Nanomachine Punches a Lethal Blow to Bugs
Published in Microbiology, Cell & Molecular Biology, and Immunology
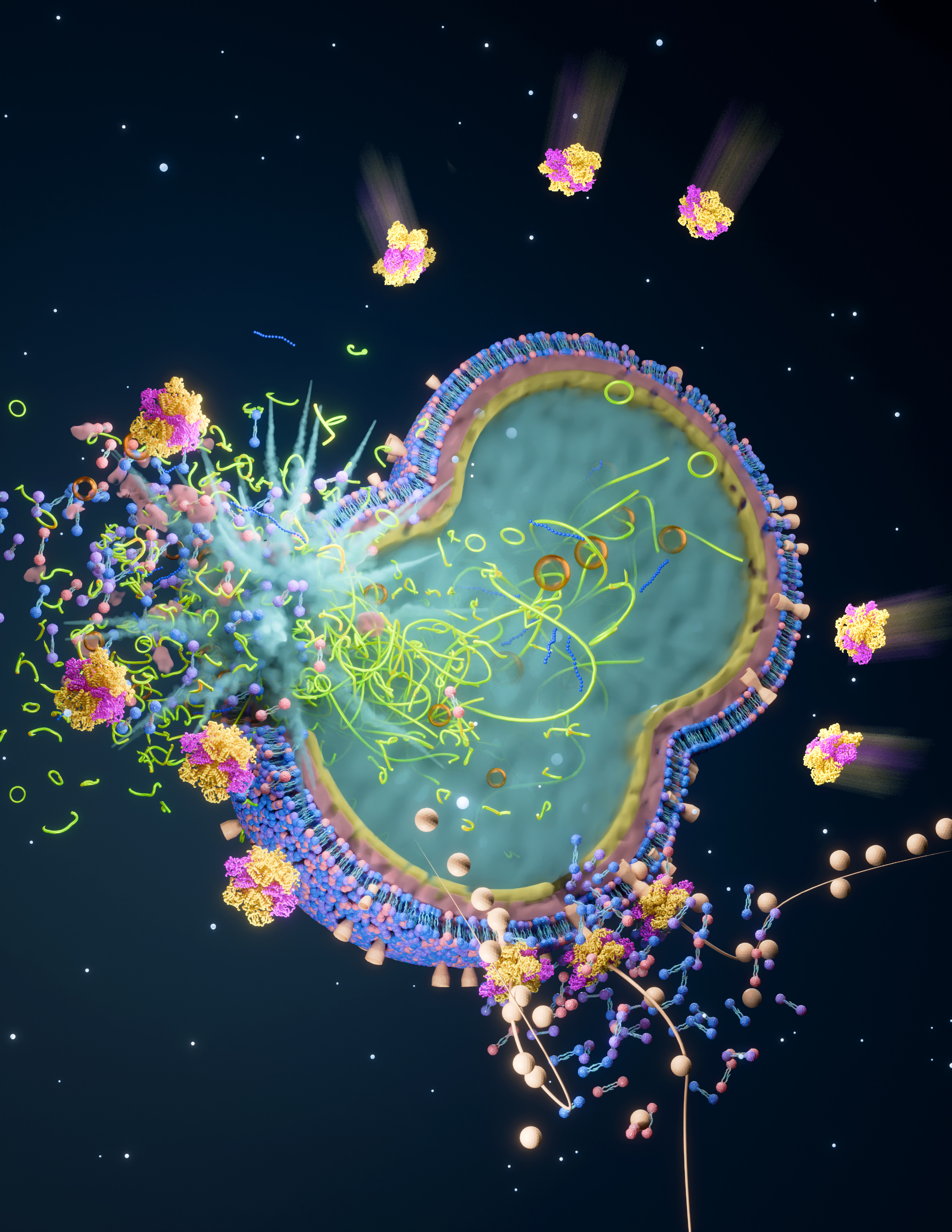
Bacteria often seek refuge inside the host cells, as it is a luscious environment for them to nourish and avoid extracellular defence mechanisms such as antimicrobial peptide or complement factor-based killing. However, there is a more sinister motive behind this strategy, which is to infiltrate the bloodstream lying deep beneath. Once the bacteria enter the vasculature, they rapidly multiply and spread to multiple organs, leading to life-threatening multi-organ failure, commonly known as sepsis. Yet, despite the constant exposure to countless pathogens, we rarely fall sick and much of the credit goes to the robust, self-regulating defence mechanisms within each of our cells. These defences are multi-layered, designed to act in a fail-safe manner, ensuring that if a pathogen evolves to bypass one barrier, others stand ready to counter it.
In recent years, cytosolic guarding factors have garnered significant attention, raising the question of what happens when bacteria escape their initial endosomal membrane and enter the cytosol of host cells in search of nutrients. A major breakthrough was identification of Guanylate-Binding Proteins (GBPs) that can either kill bacteria directly or alert the nearby cells1. Notably, the synergy between GBPs and Apolipoprotein L3, a protein with detergent-like properties, proved effective in selective pathogen killing2. Even the proteasome has re-emerged as a critical player, producing antimicrobial peptides from degradation of host proteins that disrupt specific bacterial membranes. While these findings are groundbreaking, they highlight the selective nature of these defence strategies, limiting their reach. This led us to explore ubiquitination, a more universal 'death tag' that marks various bacteria regardless of their phylogenetic background.
Previous work from our group, led by Dr. Smita Bhutda and Dr. Shruti Apte, extensively investigated how ubiquitination is involved in clearance of invading bacterial pathogens. Using Streptococcus pneumoniae, a major cause of pneumonia, sepsis and meningitis, we discovered that specific ubiquitin chain formations on bacterial surface proteins are essential for targeting these pathogens to the proteasome for degradation3. However, this led to a paradoxical scenario. For the proteasome to degrade the substrate (typically proteins), it needs to enter its narrow central pore, where the proteolytic cleavage occurs to dismantle it. However, the size of a bacterium is almost 1000 times larger. Thus, we wondered how proteasomes could achieve this. Since an entire bacterium cannot be devoured by a proteasome, we speculated that the selective bacterial surface proteins carrying the ubiquitin tag must be plucked out from the membrane and fed to the proteasomes. However, as the proteasomes lacked such extraction ability, we believed an auxiliary factor could be involved in streamlining this process. Our search led us to VCP (Valosin-Containing Protein), also known as p97, which turned out to be much more than just a feeding machine to proteasomes.
")
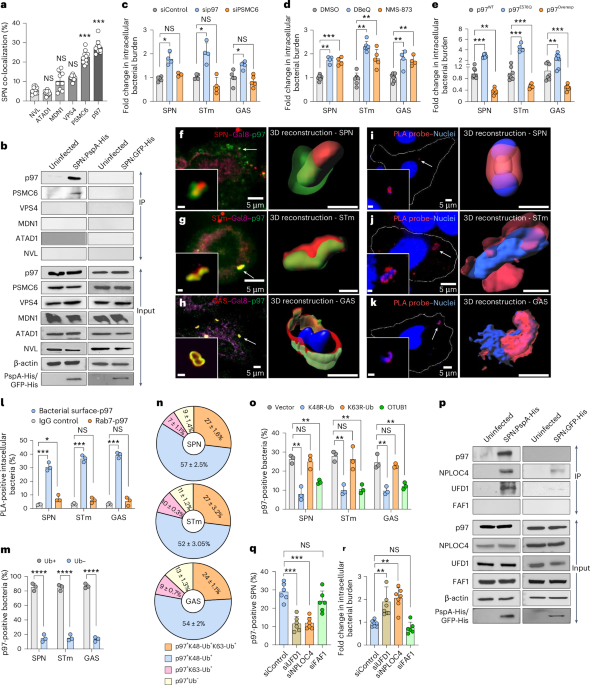
Initially, we found that VCP/p97’s targeting of the pathogen was based on four criteria: (1) the bacteria must be exposed to the cytosol, (2) display degron motif containing surface proteins, (3) be marked with K48-linked polyubiquitin chains, and (4) recruit the NPLOC4-UFD1 dimer, which helps localize p97 to the site of action. This part, involving cell biology and biochemical assays, was relatively straightforward given the expertise we developed in the lab over the years. Experiments in these lines surprisingly indicated that p97 is associated with bacterial killing. This is where the real challenge arose when we tried to understand how p97's targeting and extraction of bacterial proteins translated into actual pathogen elimination. It took us a rigorous year to establish an in-vitro reconstitution assay, where we could recapitulate the entire cellular process in test tubes by adding purified components. Standardizing this assay was a pivotal breakthrough, as it was crucial for uncovering the precise mechanism of p97’s antimicrobial action. When we saw that p97 treatment reduced bacterial viability, we were eager to examine the bacteria's morphology. We travelled from city to city seeking high-quality TEM (transmission electron microscopy) images, but the results were often disappointing and criticized for their poor quality. Just when hope was dwindling, a high-resolution TEM facility was established at our institute. With low expectations, we gave it a try, and the first image left us awestruck. Not only did it validate our hypothesis, but it also revealed intricate details beyond our imagination. Despite facing many more challenges throughout the project, overcoming these initial hurdles remains one of the most rewarding aspects of our work. Nevertheless, this led us to establish a newfound role of VCP/p97, as a bacteriolytic agent.
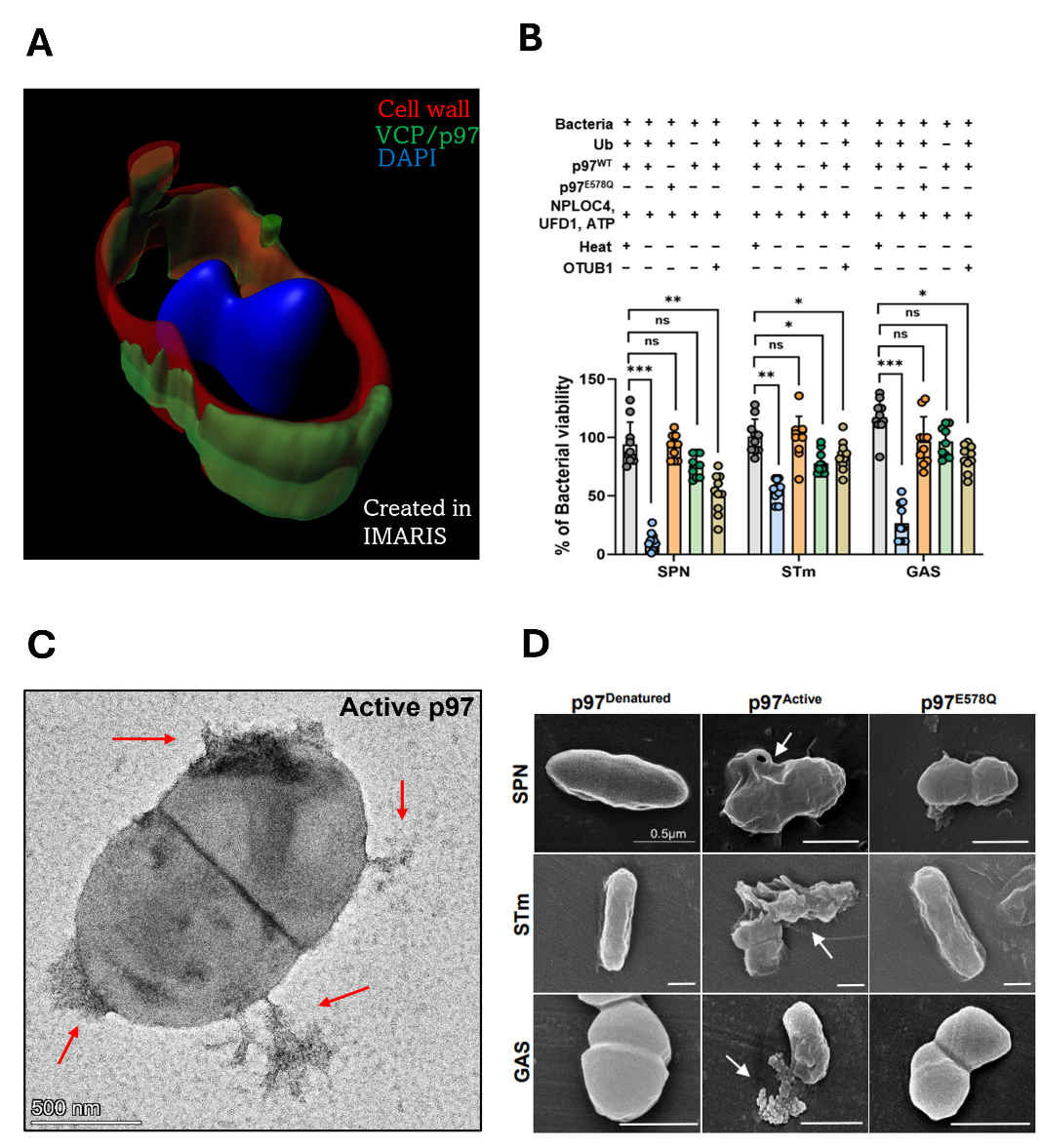
Several aspects of this work extended beyond our core expertise, but we were fortunate to have incredibly supportive and knowledgeable collaborators. Their patience and guidance were extremely valuable in helping us master the necessary techniques and accurately determine that indeed VCP/p97 could extract bacterial proteins. By employing simulations, we visualized how the removal of each protein could severely compromise the bacterial cell wall. Using optical trap, we demonstrated real time extraction of bacterial protein bound to an artificial surface. Furthermore, we demonstrated that VCP/p97 serves as a formidable barrier against Streptococcus pneumoniae, curbing its rapid proliferation in the spleen and ultimately preventing its entry into the bloodstream, thereby safeguarding the host from lethal sepsis.
By unveiling this potent, broad-spectrum antimicrobial mechanism, we open the door to novel strategies that could be exploited to outmanoeuvre rapidly evolving bacterial and non-bacterial threats, a promising step toward fortifying our defences against infectious diseases.
References:
1. Kutsch, M., Sistemich, L., Lesser, C. F., Goldberg, M. B., Herrmann, C., & Coers, J. (2020). Direct binding of polymeric GBP1 to LPS disrupts bacterial cell envelope functions. The EMBO Journal, 39(13). https://doi.org/10.15252/embj.
2. Gaudet, R. G., Zhu, S., Halder, A., Kim, B., Bradfield, C. J., Huang, S., Xu, D., Mamiñska, A., Nguyen, T. N., Lazarou, M., Karatekin, E., Gupta, K., & MacMicking, J. D. (2021). A human apolipoprotein L with detergent-like activity kills intracellular pathogens. Science, 373(6552). https://doi.org/10.1126/
3. Apte, S., Bhutda, S., Ghosh, S., Sharma, K., Barton, T. E., Dibyachintan, S., Sahay, O., Roy, S., Sinha, A. R., Adicherla, H., Rakshit, J., Tang, S., Datey, A., Santra, S., Joseph, J., Sasidharan, S., Hammerschmidt, S., Chakravortty, D., Oggioni, M. R., . . . Banerjee, A. (2023). An innate pathogen sensing strategy involving ubiquitination of bacterial surface proteins. Science Advances, 9(12). https://doi.org/10.1126/
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.

Ask the Editor - Immunology, Pathogenesis, Inflammation and Innate Immunity
Got a question for the editor about the complement system in health and disease? Ask it here!
Continue reading announcement

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in
Hello Prof. Banerjee,
your group has elucidated a major facet of innate immunity defense. It is amazing how resources are garnered, and even from bacterial sources, to mount defense. I have quoted your work in my post to explain clinical findings of injured tissue that stay free of infection in a recent article: "The Cell's autonomous response window for advanced healing".
Enrique Feoli-Tufi