The formation and stabilisation of dendritic spines, a perspective on Cdk5-dependent dynamics activated by corticotropin-releasing factor.
Published in Neuroscience
Stress significantly influences brain plasticity, triggering changes in structure and neuronal networks to adapt to environmental challenges. Corticotropin-releasing factor (CRF) is a major regulator in the stress-response. In the brain, CRF acts as an important stress-mediator by performing as a neuromodulator on neuronal activity and plasticity. In chronic stress, CRF is the critical player whose actions will lead to spine loss, loss of dendritic integrity by loss of functional excitatory synapses in the hippocampus. However, in acute stress, the impact of CRF on synaptic machinery remains inadequately understood, lacking clarity in both morphological and structural events, as well as the underlying molecular interactions.
Long-term acute CRF-dependent effects on dendritic spines:
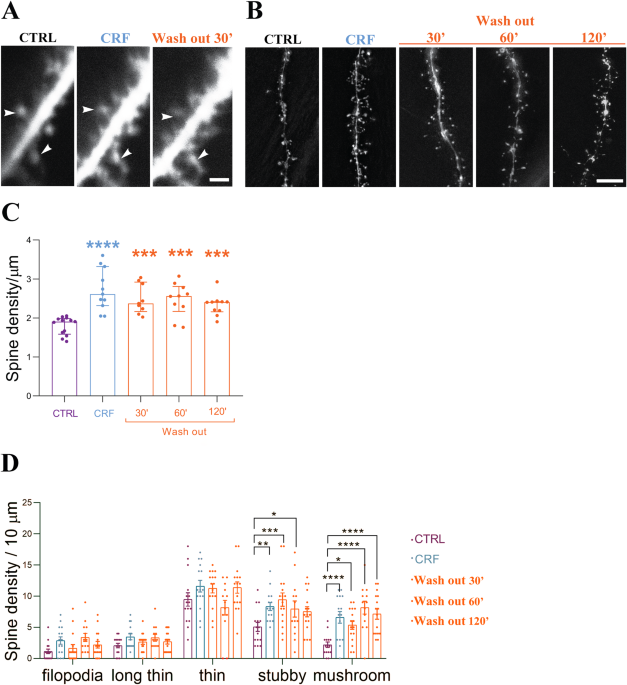
In our paper, " Cdk5-dependent rapid formation and stabilization of dendritic spines by corticotropin-releasing factor”, published in Translational Psychiatry (Vandael et al 2024), we describe the "long-term effect" of acute CRF exposure ex vivo on synaptic integrity over a time course of multiple hours (Fig. 1).
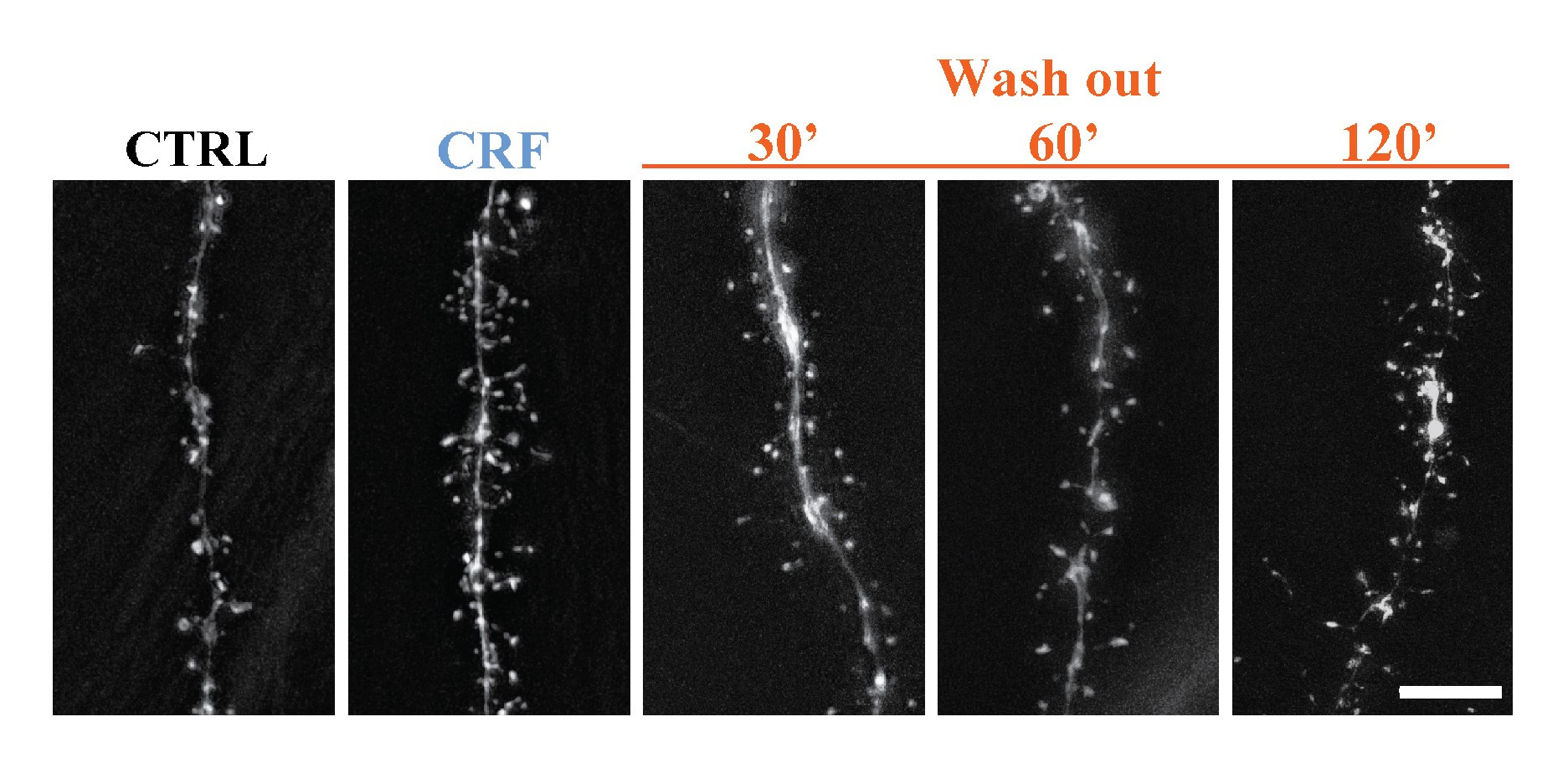
Figure 1: Acute CRF leads to acute spine augmentation in CA 1 pyramidal cells and lasts for multiple hours ex vivo.
In our previous studies, our laboratory documented the increase in spine density in the hippocampal CA1 stratum radiatum (Vandael et al. 2021) immediately following acute CRF exposure. By building on this foundation, our current investigation provides the evidence that acute CRF exposure leads to the increase in more mature mushroom spines with the employment of two-photon imaging. These spine types are more stable, and indeed even two hours after CRF exposure, a significant increase remains visible in these mature spines, indicating a stronger and more stable synaptic transmission can persist even after the removal of CRF.
In our exploration of the operational dynamics of CRF regarding the increase and stabilisation of synapses, thereby promoting enhanced synaptic transmission, we addressed two key questions. First, we aimed to identify the specific receptors involved. Subsequently, we endeavoured to elucidate the molecular mechanisms downstream of CRF-R activation.
The quest of the CRF-Rs
For addressing the first inquiry, we implemented state of the art electron microscopy techniques. By implementation of EM, we were able to confirm the presence of both CRF-R subtypes (Fig. 2) in the CA1 region of the hippocampus. Moreover, we were able to pinpoint the exact subcellular location of the different receptor subtypes within the Ca1-SR synapse.
While the availability of a reliable antibody for CRF-R1 facilitated our investigation, the situation was markedly challenging for CRF-R2. Previous studies abolished the theory of CRF-R2 presence in the hippocampus. The lack of specific antibodies for CRF-R2 further compounded the challenge. We devised a novel strategy to circumvent this obstacle. To be more specific, we conjugated a specific CRF-R2 blocker with a biotin molecule. This creative and innovative approach not only allowed us to prove CRF-R2 presence in the hippocampus using wide field microscopy but enabled us also to use immune-gold EM technique to see its subcellular localisation, like CRF-R1, at the pre-synaptic site in the CA1 hippocampal synapses. This approach provided a breakthrough in the localisation of both receptors without any potential for misinterpretation.
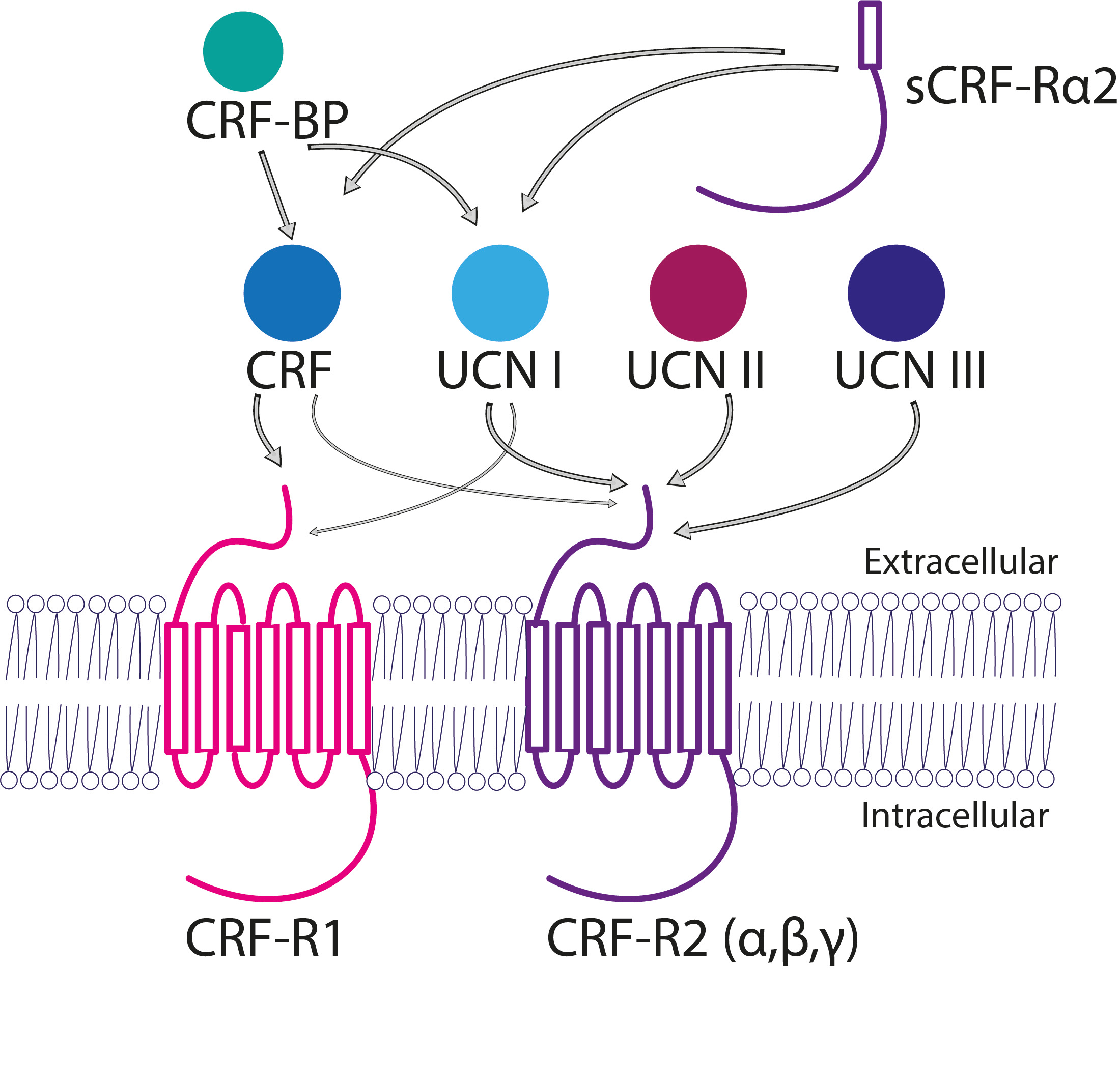
Figure 2: The CRF-peptide family. CRF and CRF-ligands can be activated by two different G-protein coupled receptors CRF-R1 and CRF-R2, respectively. CRF-ligands have different affinities for both receptors, those affinities are represented by the thickness of the arrows. CRF-R1 and/or CRF-R2 activation enables different G-protein signalling cascades. Abbreviations: corticotropin-releasing factor (CRF), urocortin (UCN), CRF receptor 1 (CRF-R1), CRF receptor 2 (CRF-R2), CRF-binding protein (CRF-BP), α-soluble isoform of CRF-R2 (sCRF-Rα2). ( Vandael and Gounko 2019).
Everything starts and ends with actin.
Due to a rapid change in the structural spine dynamics that were sustained overtime. Actin dynamics and actin rearrangements were suggested to play a role in the CRF-dependent spine augmentation and prolonged stability. In other contexts, CRF-dependent effects were reported to regulate actin-dependent signalling. But until now, downstream pathways were not clearly defined in the brain. By using a specific Cyclin-dependent kinase 5 (Cdk5)-signalling blocker Roscovitin (ROS) we aimed to demonstrate actin dynamical changes upon transient CRF exposure. By both experimental ex vivo and in vivo condition, we exposed the CA1 cells to acute CRF after this blocker ROS pre-treatment and discovered that we blocked specifically CRF-dependent spine augmentation. This suggests that a transient surge of CRF in the CA1 region of the hippocampus leads to Cdk5-dependent actin rearrangements.
Conclusion
The present study underscores the significance of the individual actions by CRF, its receptors presence and subcellular localisations in the CA1 hippocampal region. Furthermore, we demonstrate the intricate molecular mechanisms that underlie alterations in spine dynamics in acute stress. This all gives us a more profound understanding of the complex interactions between synaptic plasticity and neuropeptide-drive actions in the brain.
Follow the Topic
-
Translational Psychiatry
This journal focuses on papers that directly study psychiatric disorders and bring new discovery into clinical practice.

Your space to connect: The Psychedelics Hub
A new Communities’ space to connect, collaborate, and explore research on Psychotherapy, Clinical Psychology, and Neuroscience!
Continue reading announcementRelated Collections
With Collections, you can get published faster and increase your visibility.
Moving towards mechanism, causality and novel therapeutic interventions in translational psychiatry: focus on the microbiome-gut-brain axis
Publishing Model: Open Access
Deadline: Nov 15, 2026
From mechanism to intervention: translational psychiatry of childhood maltreatment
Publishing Model: Open Access
Deadline: Sep 30, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in