Why rats (and humans) addicted to alcohol may not respond to psilocybin treatment
Published in Neuroscience
Do psychedelics have the same effect on both healthy and diseased brains?
Psychedelic drugs are currently experiencing an unprecedented level of public and scientific interest since they first emerged in the 1960s. However, the number of unanswered questions about their mysterious effects is growing daily. Most psychedelic research today is conducted on healthy volunteers, limiting our understanding of how these drugs function in diseased brains. It’s reasonable to anticipate that they might work differently, given that each neuropsychiatric condition alters brain organization in a way that differs from a healthy individual, with its own unique homeostatic adjustments at the circuit, synaptic, and molecular levels. From the legacy studies conducted in the 1960s, many of which had questionable methodological designs and flaws, and a select few that meet today’s clinical trial standards, we know that LSD was more effective than a placebo in treating alcohol use disorder, or alcohol addiction as it’s commonly referred to. But what about animal models? Could they assist us in making well-informed decisions about patient treatment? Potentially, verifying that psychedelics act on similar brain regions in humans and lab animals, both in health and disease, could significantly broaden our ability to study these drugs using invasive methods that are otherwise impossible to apply to humans. One potential characteristic of psychedelic drug action is a decrease in connectivity within brain regions that make up the default mode network, a finding that has been confirmed in numerous studies with healthy volunteers. In our study, we tackled both of these issues: we sought to find translational evidence for the role of the DMN in psilocybin’s acute effects in rats, and questioned how the severity of alcohol addiction might influence the overall response to psilocybin.
What was our approach?
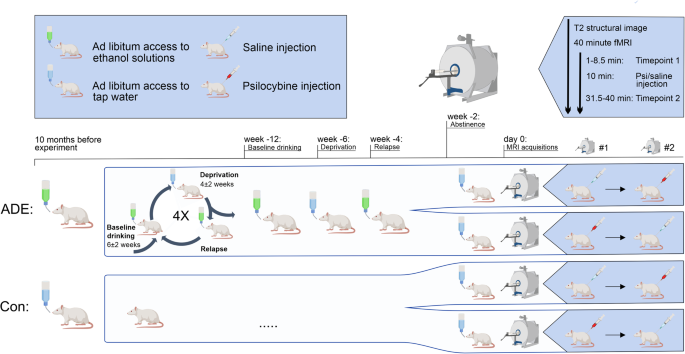
We used 25 female Wistar rats from our own breeding line as model organisms. Fifteen of these rats underwent 12 months of intermittent cycles of voluntary alcohol consumption and deprivation, while the remaining 10 served as controls. Each rat in the alcohol group had a sort of “minibar” in their home cage, where they could freely choose to drink from water, 5%, 10%, and 20% ethanol. After an initial 3 months of drinking, we removed the alcohol-containing bottles from the cages, initiating a 2-week deprivation period. Following this, the bottles were returned and their daily consumption was measured. These cycles of drinking and deprivation continued for 12 months. Each time after the deprivation, the rats would consume significantly more alcohol than they did in the week before the abstinence, a measure of relapse known as the Alcohol Deprivation Effect (ADE). The severity of relapse was defined as the percentage change in alcohol consumption during relapse compared to the baseline - the week before the abstinence. Before undergoing MRI scanning, the rats were subjected to one final 2-week deprivation period. Scans were taken using a Bruker BioSpec 9.4 T MR-scanner under combined isoflurane/medetomidine anesthesia, a common gold standard for resting-state fMRI in rats. Initially, we took a structural MRI scan to create a 3D anatomical image of the rat brain. We then took a functional MRI scan to serve as a baseline, injected the drug or placebo, and performed another functional scan at the time when the peak effects of the drug should have been reached. Each rat from the alcohol and control groups underwent two scan sessions, one with a placebo and one with psilocybin (a crossover design). We analyzed three main parameters: Global Brain Connectivity (GBC) - how each voxel is connected with every other voxel in the brain; Network-Based Statistic (NBS) - how brain regions consisting of groups of voxels are interconnected; and local Default Mode Network (DMN) connectivity - for every placebo and psilocybin condition. Finally, we compared the functional scans of addicted rats with varying degrees of relapse severity to see if they would influence the mode of action of psilocybin.
What did we discover?
We noticed a significant decrease in global connectivity throughout the brain due to the immediate effects of psilocybin. The regions primarily affected included cortical areas such as the insular and cingulate cortices, auditory and visual areas, the association cortex, and the hippocampus. When we focused our analysis on specific regions of interest, we identified a network with reduced connectivity that included regions similar to those observed with global brain connectivity (GBC). However, we also found that another circuit, specifically the dorsal raphe nucleus (DRN) and hypothalamus’ connections to various cortical areas, was hyperconnected. The DRN is the source of all serotonergic neurons in the brain and regulates cortical reorganization among different sensory and effector systems. This observed hyperconnectivity could indicate increased neuroplasticity, which is known to be triggered by psychedelics.
When we specifically examined the default mode network (DMN) under the immediate influence of psilocybin, we noticed a significantly reduced connectivity pattern that affected 9 out of the 12 regions that comprise the DMN. To the best of our knowledge, this is the first direct replication of human resting-state fMRI with psilocybin in rats. This suggests that despite approximately 75 million years of evolutionary divergence between humans and rodents, the way psilocybin impacts our brains is remarkably similar, a finding we were able to confirm using fMRI.
But there’s more. When we examined global and DMN-specific brain connectivity in relation to the severity of relapse, we found an inverse correlation: the stronger the relapse, the less decrease in connectivity was observed under psilocybin. In other words, the brain was more “resistant” to its immediate effects, both globally and at the DMN levels. This suggests that, at least in rats, the severity of alcohol addiction may impede the potential therapeutic effects of psilocybin and other psychedelics, particularly if they are administered at a standard dose. One possible explanation could be anaplasticity, a gradual loss of neuronal and molecular plasticity following drug exposure in the addicted brain, resulting in rigid behavior and failure to respond to the standard dose of a psychedelic. It is also known that alcohol-dependent rats have decreased expression levels of mGluR2, a receptor that forms heteromeric complexes with 5-HT2A, the primary target of psychedelics. Its loss in the prefrontal cortex is likely to alter the effects of psilocybin in these rats.
In conclusion, we provided the first translational evidence of preserved functional connectivity changes under psilocybin between rats and humans, which is vital for advancing preclinical research on psychedelics. Furthermore, we discovered that in a rat model of alcohol relapse, a stronger relapse is associated with weaker psilocybin effects. This suggests that future clinical trials may need to adjust psychedelic dosages based on the severity of an individual patient’s condition. There is still a long journey ahead to understand the mystery of psychedelic action in the brain, and we are pleased to have moved this exciting journey one step further.
Follow the Topic
-
Translational Psychiatry
This journal focuses on papers that directly study psychiatric disorders and bring new discovery into clinical practice.

Your space to connect: The Psychedelics Hub
A new Communities’ space to connect, collaborate, and explore research on Psychotherapy, Clinical Psychology, and Neuroscience!
Continue reading announcementRelated Collections
With Collections, you can get published faster and increase your visibility.
Moving towards mechanism, causality and novel therapeutic interventions in translational psychiatry: focus on the microbiome-gut-brain axis
Publishing Model: Open Access
Deadline: Nov 15, 2026
From mechanism to intervention: translational psychiatry of childhood maltreatment
Publishing Model: Open Access
Deadline: Sep 30, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in