Capturing cellular rearrangements in developing sensory epithelia
Published in Ecology & Evolution
Vertebrates possess highly developed sense organs, responsible for detecting information about different environments and converting extracellular stimuli into electrical signals, which are mediated by sensory epithelia. These epithelia have been observed to exhibit mosaic patterns through continuous intercalations of sensory and supporting cells (SCs). Dr. Hideru Togashi (Kobe University, Japan) and his team hypothesized that such pattern formation is due to varying adhesion strengths between participating cells. This idea was supported by their numerical results generated via vertex dynamics of polygonal cells. However, it was found that frequent occurrence of cell intercalations precludes the application of vertex dynamics to support this hypothesis through simulations - this prompted our collaboration with Dr. Hideru Togashi and Dr. Hideki Murakawa (Ryukoku University, Japan).
To analyze the effect of such factors in tissue morphogenesis, we assume, as in the vertex dynamics model, that tissue evolves via a succession of quasi-equilibrium states, that is, cell shapes are described by their instantaneous state of lowest energy. To be precise, we treat cellular rearrangements as the gradient flow of the weighted surface energy constrained by each cell's prescribed volume. Here the weights are derived from measured values such as cell-cell adhesion or interfacial tension. In addition, we assume that no apoptosis occurs during the evolution. This simplification can, however, be made more realistic by allowing varying cell volumes.
We proposed a level set-based computational framework for the numerical realization of this model. Our initial approach was to embed the geometry of an aggregate of n cells in an (n-1)-vector field and employ our previous work on multiphase volume-preserving interface motions, which involves approximating the minimizer via gradient descent method. Unfortunately, this scheme was computationally costly; and a much faster approach was necessary. With this, we turned to Esedoglu and Otto's algorithm. Here the geometry of each cell is implicitly represented as a level set of some function. The main idea is to solve heat equations via fast Fourier transform and to extract level sets of their solutions to determine the geometry of the cell aggregate in the next time step. In order to apply this scheme, two issues need to be addressed: volumes of cells have to be preserved or controlled, and cells have to be prevented from splitting. We implemented volume control via auction dynamics, which in essence finds suitable contour heights by simulating an auction. The second issue relates to the absence of any scheme in the original algorithm that would ensure local cell connectivity. It may happen that a cell splits into two parts and one part suddenly appears in a distant part of the aggregate. To avoid such behavior, we proposed a localization scheme on the bidding process in auction dynamics.
Our computational analysis via a series of numerical tests revealed that the proposed method converges even across topology changes; yields better results in comparison to the vertex dynamics algorithm at the price of higher computational cost; generates expected results for standard morphogenetic benchmarks, namely cell sorting, mixing and formation of checkerboard and football patterns; and handles simulations in higher dimensions without major technical difficulties or theoretical complications. It is worthwhile noting that these cell morphologies involve curved cell-cell junctions, largely different sizes of participating cells, and frequent topology changes. Even then, our proposed level set approach can easily handle such complexity, not only because it allows for a wide range of geometrical patterns, but also because it implicitly satisfies the precise contact angles at tricellular junctions, which is indispensable for theoretically correct realization of cellular intercalation processes.
Now that the numerical method is established, we circle back to what started this research project - to computationally reproduce observed cellular patterns in developing sensory systems, in particular, olfactory (OE) and auditory epithelia (AE).
(Reproduced from Figure 3, Mohammad et al. 2022)
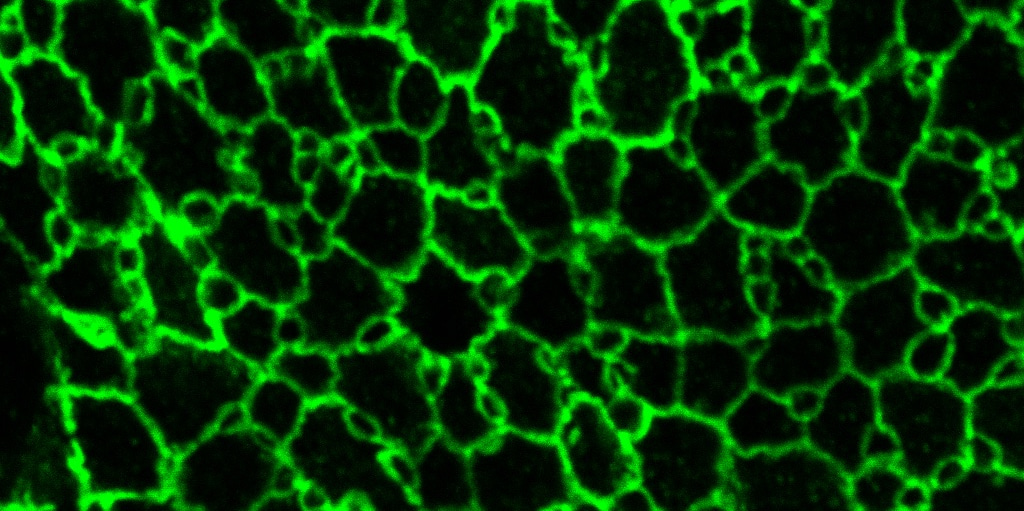
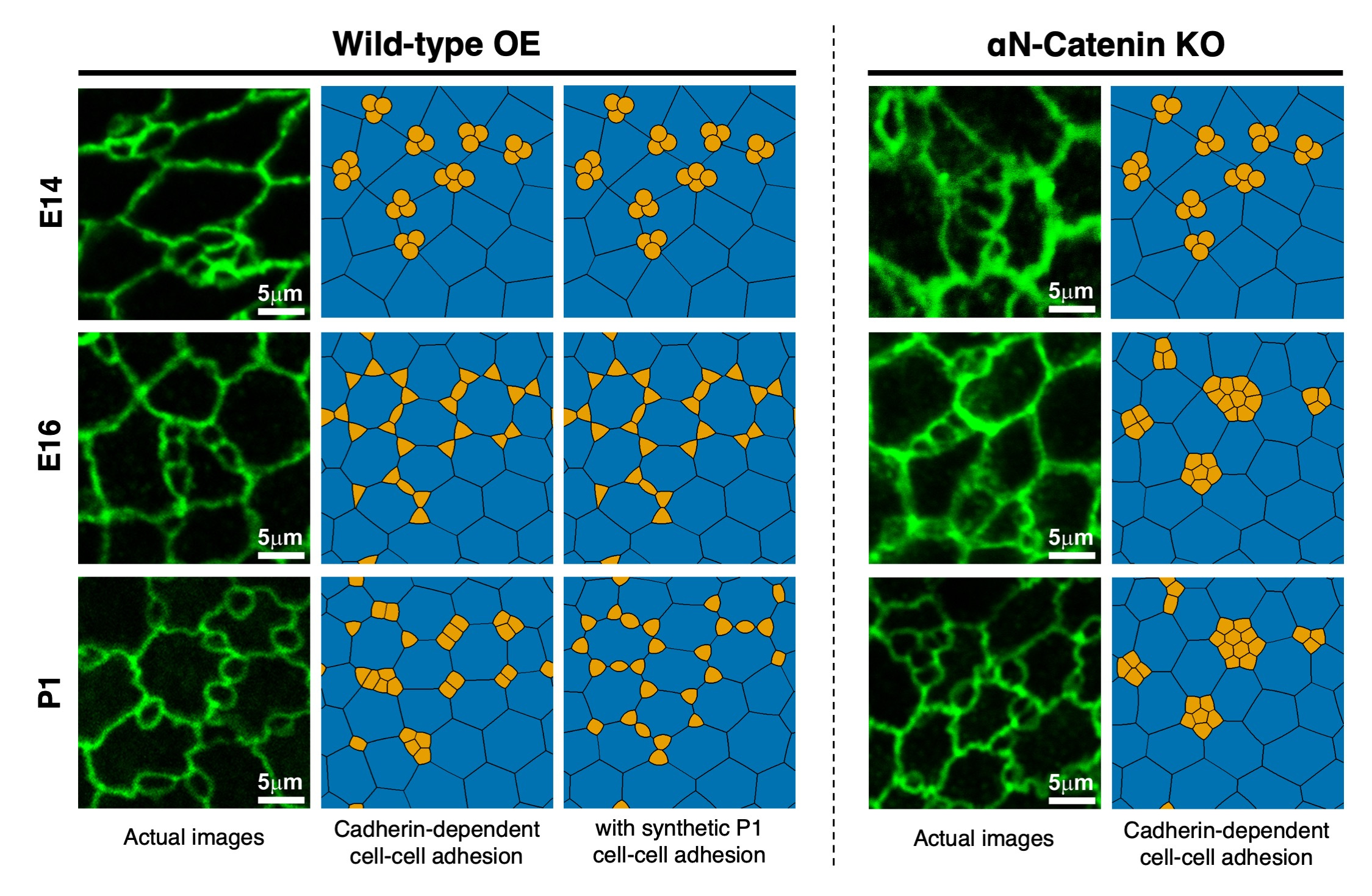
The OE, which is located inside the nasal cavity, is a specialized sensory epithelium involved in odor perception. When the luminal surface of the OE is observed from the apical side, ciliated olfactory cells (OCs) and several types of SCs arrange in a unique mosaic pattern accompanied by cellular rearrangements from embryonic day 14 (E14) to postnatal day 1 (P1) as shown in the Figure 1. To estimate cell-cell adhesion strength, we use the distributional patterns of experimentally measured β-catenin intensities, from wild-type (WT) and αN-catenin KO mice, since in this case, cadherin-dependent affinity is the major component of cell-cell adhesion. With this, we employed our proposed algorithm to simulate OE cellular rearrangements in WT and αN-catenin KO mice.
Comparing simulation results to biological experiments, Figure 1 shows that our model was able to capture overall cellular rearrangements in the embryonic stage. In particular for WT, OCs cluster at the tricellular SC-SC junctions at E14; then from E16, OCs separate and move along SC-SC junctions. The only noticeable difference is that OCs at tricellular junctions are not as round as in the experimental results. Moreover, postnatal simulations indicate that β-catenin and, thus, cadherin-dependent cell-cell adhesion, is not the only contributing factor for cellular rearrangement in OE. This discrepancy is caused by an insufficiency in the model parameters, rather than by the proposed numerical method or the model itself. To verify this, we choose cell-cell adhesion strengths that imitate the hypothesized prominent activity of cytoskeleton within OCs in postnatal stages. The resulting WT cellular rearrangements exhibit a clear agreement with the actual P1 pattern: OCs are separated one from another and are located either at tricellular junctions or in the middle of SC-SC junctions. We conclude that our proposed algorithm is able to simulate complex cell behaviors in evolving OE across all stages until P1, while the physical model based on β-catenin mediated adhesion captures well the phenomenon only in embryonic stages.
(Reproduced from Figure 5, Mohammad et al. 2022)
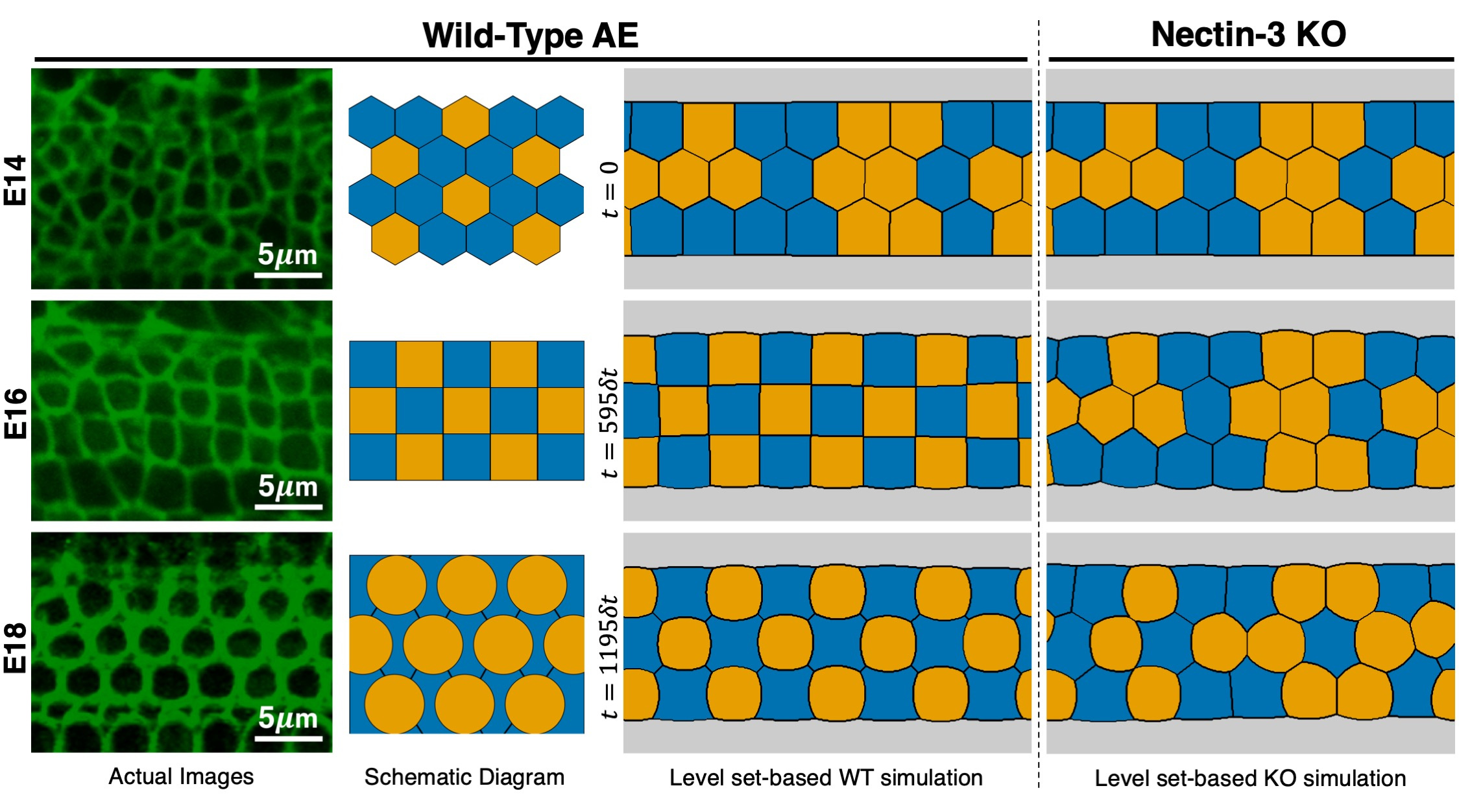
The AE, which is responsible for hearing, is composed of mechanosensory hair cells (HCs) and SCs that rearrange to form a checkerboard pattern from E14 to E16. At E18, HCs arrange in ordered rows, and each HC is separated from one another by a SC, forming an alternating mosaic pattern. During embryonic development, heterophilic molecular interaction between nectin-1 on HCs and nectin-3 on SCs is much stronger than the homophilic interactions of nectins between the same type of cells. This biased cell-cell adhesion in WT is responsible for the checkerboard assembly of cells. Consequently, the absence of nectin-3 in KO mice eliminated this bias resulting in an overall disruption of the checkerboard pattern. Our level set-based simulations revealed that the expected WT and nectin-3 KO patterns from E14 to E18 stages can be reproduced solely by designing the interfacial tensions (see Figure 2). For WT, we start with equal interfacial tensions at E14, then decrease only the SC-HC tension, which qualitatively follows the change in adhesion strengths measured experimentally in terms of β-catenin intensities at E16. Moreover, pattern formation after E16 cannot be explained only by adhesion - the change in hair cell morphology from square to circular is thought to be caused by pronounced cytoskeletal dynamics within hair cells, leading to increased hair cell stiffness. To express this in our model, we evolve interfacial tensions so that HC-SC tension is less than HC-HC, but greater than SC-SC. For nectin-3 KO, the interfacial tensions are set so that the strong adhesion between HCs and SCs is inhibited at E16. These simulations indicate the importance of transitory formation of checkerboard pattern at E16 for robust attainment of mosaic distribution of HCs and SCs. Although the nature of the relationship between interfacial tension and adhesion, contractility, stiffness, internal pressure, etc., is still unknown, our results show the relevant potential of our level set-based approach as an effective tool for investigating this relationship.
(Reproduced from Supplementary Figure 16, Mohammad et al. 2022)
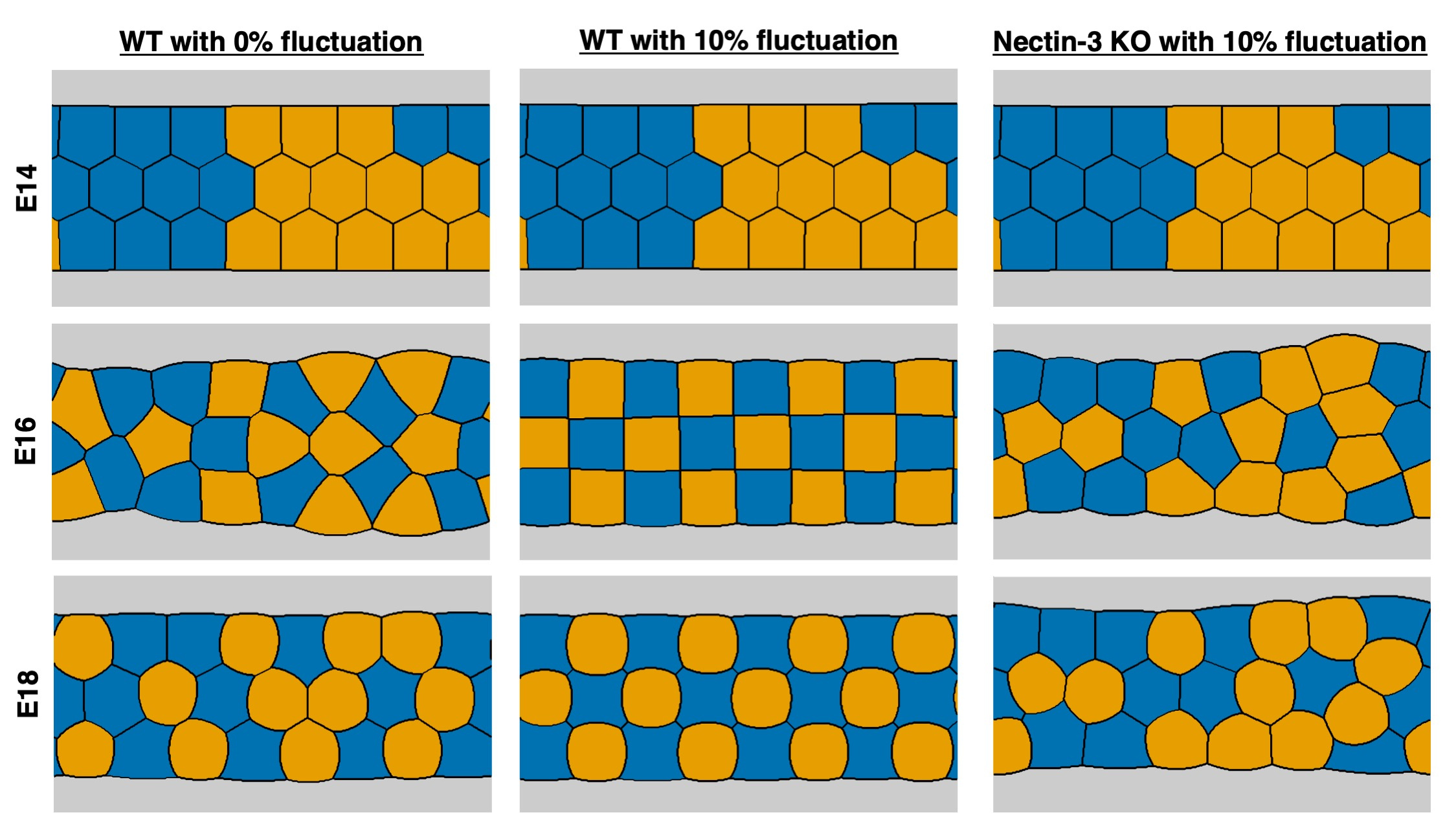
Finally, we note that simulation results may depend on the initial configuration as shown in Figure 3. Under this setup, perfect checkerboard pattern at E16 and the expected final pattern at E18 were not reached. This dependence, however, is eliminated by including fluctuations in the model since various factors in living tissues, e.g., cellular fluctuations, mechanical forces, induce relocation of cells. This simulation indicated that independent of the initial configuration, the combination of the fluctuations and the affinity between HCs and SCs at E16 lead to a regular checkerboard pattern and subsequently to the desired highly organized pattern at E18. Elucidation of the mechanism of pattern formation requires further investigation and our proposed level set-based scheme can be a reliable tool for pursuing this research.
In sum, applying our proposed level set-based method to the analysis of the underlying mechanism of pattern formation in sensory epithelia, led us to discover that indeed, differential interfacial tension plays a decisive role. By simulating the epithelial formation based solely on interfacial tensions, we were able to reproduce the evolution of cellular patterns observed in experiments. Such finding was not possible until now because established methods either were not able to capture complex curved shapes of cells with largely differing sizes and/or were not sufficiently mathematically accurate to grasp the frequent and delicate topology changes, such as intercalations, that are at the core of the patterning process.
If you enjoyed this read, feel free to visit our open-access paper at: https://www.nature.com/articles/s42003-022-03174-6
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Mechanistic insights into human host and microbiome interactions
Publishing Model: Open Access
Deadline: May 31, 2026
Advances in neurodegenerative diseases
Publishing Model: Hybrid
Deadline: Jun 30, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in