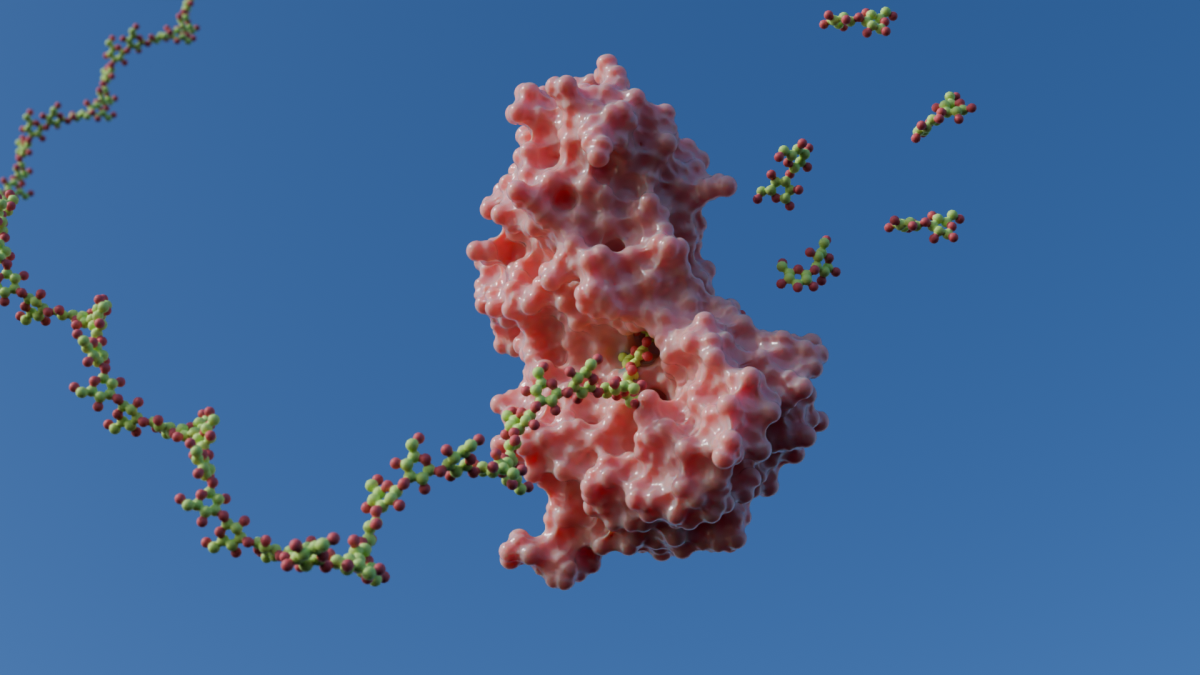

β(1,3)-glucans are a class of carbohydrates found across different kingdoms of life, being fundamental components of fungi, algae, plants and bacteria cell walls. The β(1,3)-glucans depolymerization process has attracted increasing biotechnological interest, especially in health and food industry. For a long time, β(1,3)-glucan deconstruction has been traditionally viewed through the lens of classical endo-acting enzymes. These enzymes are able to bind to the polymeric chain and promote hydrolysis internally, releasing the products upon dissociation. Then, the enzyme binds to another carbohydrate chain or fragment to begin a new catalytic cycle.
Recently, we recovered an unconventional member of the Glycoside Hydrolase family 158 (GH158) from a metagenomic library of soil samples covered with sugarcane bagasse. This enzyme, expected to act as a typical endo-β(1,3)-glucanase, generated a disaccharide, the laminaribiose (L2), as the predominant product. This was an indicative that the enzyme could perform a processive mechanism for catalysis, instead of being a classical endo-glucanase. Processive catalysis is a mechanism where the enzyme binds to a polymeric substrate (like beta-1,3 glucans) once and performs multiple consecutive catalytic cycles before dissociating from the chain. This type of mechanism is known for many biological systems and is adopted by several enzymes in Nature, such as DNA polymerases and cellobiohydrolases, an efficient model for the cleavage of cellulose. This observation became the starting point of our recent work in Nature Communications, “Conformational gating mechanism for processive catalysis of β-(1,3)-glucans'' https://www.nature.com/articles/s41467-026-71224-2, in which we combined structural biology, enzymology, and molecular simulations across multiple levels of theory.
The structural tour de force to uncover a new mechanism
We collected an extensive series of crystallographic datasets, including cryogenic and room-temperature conditions, at Sirius, the 4th-generation synchrotron located in Campinas, Brazil. In total, we solved 19 structures of the enzyme under different conditions, including several complexes with β-glucan fragments. These crystallographic structures revealed different arrangements of substrates and products within the enzyme active site.
Notably, in all productive structures, the product extended no further than the +2 subsite, corroborating the observed cleavage pattern. Furthermore, the structures showed that the active site was not an open cleft. Instead, upon substrate binding, the enzyme adopted a tunnel-like conformation that enclosed the substrate, trapping the glucan chain inside the catalytic site. A tunnel architecture is a hallmark of many processive enzymes that move along polymeric chains. It was the first time that this particular active-site architecture was verified for an enzyme involved in β(1,3)-glucan degradation in this particular way.
Revealing the dynamic behavior of the active-site tunnel and the stages of processivity
To investigate the role of tunnel architecture dynamics in catalysis, we conducted force-field-based metadynamics simulations. The simulations showed that, in the absence of substrate, the tunnel-forming loops spontaneously open. However, when the enzyme was bound to the substrate, the closed tunnel-like conformation was stabilized.
Time-course experiments by mass spectrometry, allowed us to follow product formation comparing the wild-type enzyme and mutants where the tunnel was disrupted. These experiments showed that when the tunnel-stabilizing interactions were disturbed, the product profile changed. Instead of the accumulation of disaccharide, the enzyme released a broader mixture of oligosaccharides, resembling a classical endo-dissociative enzyme.
Microsecond-timescale simulations of GH158Pro complexed with the remaining substrate chain and the product (L2), mimicking the structural organization after cleavage, then helped explain this behavior at the molecular level. The active-site tunnel undergoes conformational changes that allow the glucan chain to slide forward into a new catalytic position, accompanied by a reorganization of the active-site environment. This movement was correlated with a conformational switch controlled by a salt bridge between Arg104 and Asp382. Structurally, this interaction stabilizes the closed tunnel conformation. Functionally, disrupting this salt bridge changed how the enzyme processed the substrate. In other words, disturbing the gate changed the way the enzyme “moved” along the glucan chain.
Looking at the chemistry of the processive catalysis
We also seek to understand the chemical basis of the reaction catalyzed. QM/MM metadynamics simulations were employed to reveal the conformational itinerary (i.e. the conformational changes suffered by the substrate reactive sugar to enable catalysis). We found an unusual itinerary, starting from a relaxed 4C1 conformation that moves toward a 4H3-like transition state, and then returns to 4C1 after cleavage. Cyclic itineraries, like the one found, have been associated with exo-acting enzymes. Its presence in a processive β(1,3)-glucanase with a dynamic tunnel suggests that it could be more widespread than previously thought. The possible reason for this finding is that the leaving group remains relatively exposed to the solvent at the end of the short tunnel, which may reduce the need for strong sugar distortion before catalysis, since more degrees of freedom of the substrate is previously allowed.
Why do these discoveries matter?
Together, structural snapshots, enzyme assays and simulations revealed that the discovered enzyme does not rely on a permanently closed tunnel, but on a conformational gating. The active site opens and closes in coordination with product release and substrate translocation. Broadly, this study expands the concept of processive catalysis across one of the main polysaccharide classes found in nature. This discovery also carries biotechnological relevance, since β(1,3)-glucans have broad applications ranging from sustainable biorefineries to nutraceuticals and food additives. Understanding how nature evolved to process these complex substrates may help to guide future enzyme engineering efforts.
-glucans cleavage. The key conformational change is associated with disruption of the salt bridge and opening of the tunnel. This transition lowers the barrier for product release after cleavage and helps explain the progressive accumulation of reaction products over time.")
Graphical summary of the proposed processive mechanism for β(1,3)-glucans cleavage. The key conformational change is associated with disruption of the salt bridge and opening of the tunnel. This transition lowers the barrier for product release after cleavage and helps explain the progressive accumulation of reaction products over time.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in