How bacteria resist the oldest class of synthetic antibiotics
We discovered the molecular mechanism underpinning sulfonamide resistance by the Sul enzymes and open the door to modifying these drugs to overcome resistance.
Published in Microbiology

The first synthetic antibiotics
Sulfonamide antibiotics were first synthesized in the 1930's and today they are used to treat urinary tract infections, meningitis, pneumonia, ear infections and other indications caused by Gram-positive and Gram-negative pathogens.

Sulfonamides target the bacterial folate pathway, an essential pathway for the synthesis of tetrahydrofolate. Sulfonamides bind to and competitively inhibit the product of the folP gene, dihydropteroate synthase (DHPS) by acting as structural analogs of one of the co-substrates of DHPS, p-aminobenzoic acid (pABA) (see below).

Transferable resistance to sulfonamides
Transferable resistance to sulfonamides was discovered in the 1950s and 1960s and then delineated to the sul genes - sul1, sul2, sul3 and sul4. These genes encode dihydropteroate synthase enzymes that are divergent in sequence from DHPS (~30% identical).
Since their initial discovery, the sul genes have disseminated broadly in pathogenic species via their frequent occurrence on plasmids containing multiple antibiotic resistance genes. Today, sul1 is one of the most widely spread antibiotic resistance genes worldwide.
Our motivations and research question
We are interested in the 3D structure, function of molecular mechanisms of proteins that confer antibiotic resistance. We realized that the 3D structure and molecular mechanism of the Sul enzymes have never been characterized, even decades after the discovery of sul1.
We were motivated to answer the question: how are the Sul enzymes insensitive to sulfonamide inhibition, but nonetheless function as dihydropteroate synthase enzymes?
Enzymology of recombinant DHPS and Sul enzymes clearly reflects drug susceptibility
We recombinantly purified the E. coli DHPS enzyme (EcDHPS) and the Sul1, Sul2 and Sul3 enzymes and developed an in vitro activity assay to evaluate their reactions. Our kinetics analysis showed unambiguously that EcDHPS happily uses either pABA or SMX as substrates, but the Sul enzymes vastly prefer pABA over SMX. We also showed that SMX could inhibit the EcDHPS reaction but not the Sul reaction (at least not very well, Ki values were more than 74-times higher).
These results clearly showed that the Sul enzymes somehow select for pABA binding over SMX.
Crystal structures of the Sul enzymes
Next we crystallized all of the Sul enzymes and were fortunate enough to determine multiple ligand-bound structures, including a fascinating structure of the Sul3 enzyme in complex with a reaction intermediate. This intermediate is the pterin ring left after cleavage of DHPP by the enzyme, pyrophosphate was still bound in the active site, and the incoming pABA was also present - incredible! These structures showed that multiple loops in the active site are mobile and organize themselves to perform catalysis.
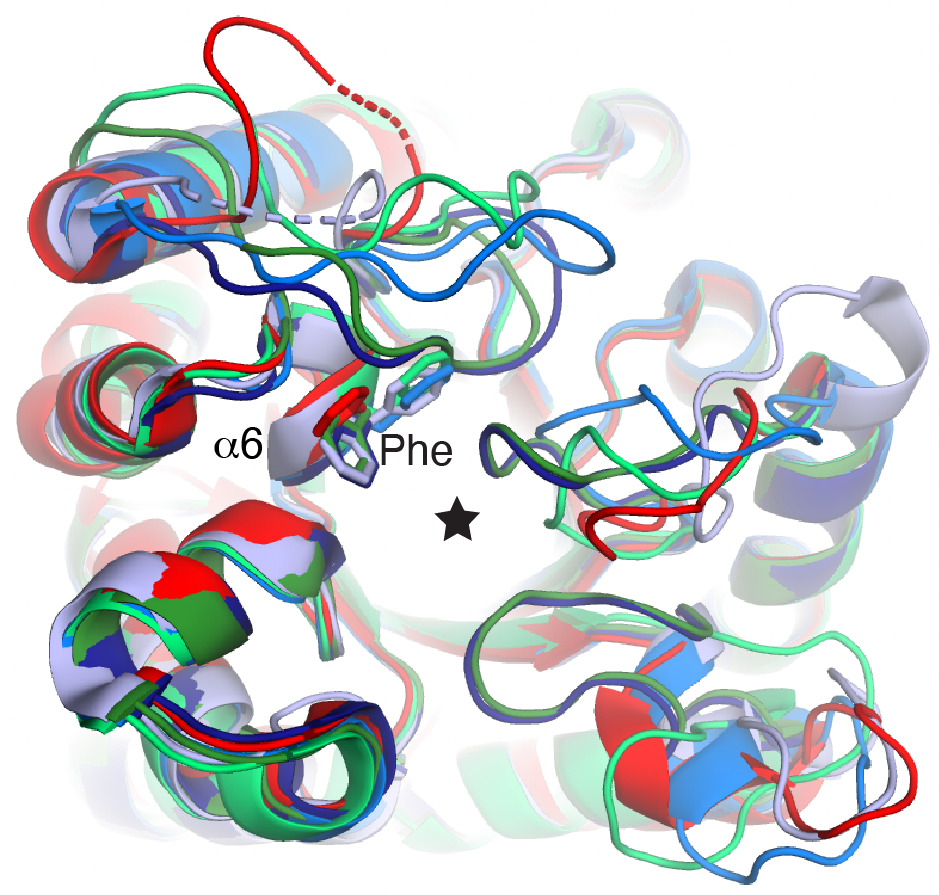
The crystal structures also showed that each of the Sul enzymes contains a phenylalanine (Phe) residue right on top of the pABA molecule (below left, in red). But none of DHPS enzymes that we analyzed in the Protein Data Bank contained this Phe residue, while the rest of their active sites and positioning of ligands was pretty much exactly like the Sul enzymes. This was an "ah-ha" moment - it made total sense that the Phe residue blocks sulfonamides from binding, but allows pABA to bind.


When we mutated this Phe residue to a glycine (Gly) in Sul1 and Sul3, this transformed their in vitro characteristics as if they were sulfonamide-susceptible DHPS enzymes.
In cellulo evaluation of sulfonamide resistance by the Sul enzymes
We wanted to quantify the degree of resistance conferred by the Sul enzymes and variants thereof that were designed based on our structural analysis. We generated a new folP knockout strain of E. coli, to remove any background effect of the DHPS/FolP enzyme, then expressed the sul genes on a plasmid. This showed clearly that the Sul enzymes conferred high-level, broad-spectrum sulfonamide resistance. Expression of the Sul Phe->Gly variants nearly completely abrogated resistance, confirming the role of this residue in selecting against drug binding.
Lab evolution of a sulfonamide-resistant DHPS enzyme that looks just like the Sul enzymes
The last experiment we performed was to see if we could evolve EcDHPS to be sulfonamide-resistant, which has been done before, but we wanted to capture modifications in its active site that resemble the Sul enzymes. We evolved E. coli in the presence of sulfonamides and trapped a mutation in folP that encodes a two-amino acid insertion in the DHPS/FolP active site - a Phe and a Gly residue (see the sequence alignment below). We then crystallized this enzyme and were shocked to observe that this inserted Phe is positioned very closely to where this Phe exists naturally in the Sul enzymes! This is shown below and red arrows show the insertion. This was another indication that this type of modification in the DHPS active site can confer drug resistance.

Going forward: searching for more sulfonamide-resistance enzymes and re-designing the drugs
With this knowledge, where do we go from here?
Firstly, we can improve our surveillance for this form of sulfonamide resistance. Since it is clear that we found a new area of the active site of dihydropteroate synthase enzymes that can vary in sequence to confer resistance, either naturally or by selective pressure, we are searching for new sulfonamide resistant DHPS/Sul enzymes with this motif. Maybe there are more "Sul" enzymes out there, and we already found some E. coli isolates in the NCBI Genbank that have a Phe-Gly insertion in their putative FolP enzymes.
There is also the Sul4 enzyme, the most recently discovered Sul enzyme, which we did not study here but are now characterizing.
Importantly, we believe these molecular and structural details can give ideas about the chemical liabilities in the sulfonamide compounds and could suggest ways to modify their structures to evade this type of resistance.
After nearly 90 years since their first synthesis, perhaps we can "correct" and improve these compounds and add new tools to our antimicrobial arsenal.
Senior Research Associate, University of Toronto
Follow the Topic
Microbiology
Life Sciences > Biological Sciences > Microbiology
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
A selection of recent articles that highlight issues relevant to the treatment of neurological and psychiatric disorders in women.
Publishing Model: Hybrid
Deadline: Ongoing
Biosensing
With this cross-journal Collection, the editors of Communications Biology, Nature Biomedical Engineering, Nature Sensors, Nature Communications, and Scientific Reports welcome the submission of primary research Articles focusing on the development of engineered biosensing devices with the potential to be applied in biomedical research and in the management of disease conditions.
Publishing Model: Hybrid
Deadline: Sep 30, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in
Great work. Very interesting. Thank you for sharing.