Linking tissue injury to neural precursor reprogramming in gliomagenesis
Published in Cancer
It has been recognized for many decades that primary brain tumours, perhaps none more than glioblastoma (GBM), consist of mixtures of primitive and more differentiated cell types within the malignant population. As well, these tumours are also found to contain a myriad of microenvironmental cell types. These nonmalignant cells include microglia, vascular cells, and other normal neural cell types such as neurons and glia. In addition, in GBM, there are other cell types, such as those that express mesenchymal transcriptional states, that are unique to the malignant process and do not have normal brain counterparts.
In this new study, we show that GBM-genesis is closely intertwined with an injury process, involving the emergence of a neural-crest transcriptional identity in oncogene-initiated neural precursor cells, which is also seen in normal neural precursors after a wound to the brain. These cells may be drivers of microenvironmental changes that contribute to tumour growth, and may be drivers of the extreme GBM cellular diversity.
Our laboratory has been studying endpoint GBMs for the past 20 years using a conceptual and experimental lens of stem cell and developmental biology, mostly by studying the in vivo function of fresh or briefly cultured human tumour cells from patient surgical samples, pointing at GBM growth reflecting a developmental hierarchy1,2. Questions always remained about why we see cells with mesenchymal transcriptional states in a neural parenchymal neoplasm. With colleagues at University of Toronto (Trevor Pugh, Gary Bader, Mathieu Lupien) and Calgary (Sam Weiss) we previously found that populations of GBM cells that had transcriptomes reminiscent of neural development were mixed with cells with transcriptomes reflecting an injury process3,4.
Clinical observations in case reports have observed GBM arising in regions of brain injury by trauma or stroke, but wider epidemiological data about the role of injury has been more equivocal. The role of tissue injury in cancer, although an age-old concept, has been receiving increasing attention in the literature, such as in pancreatic (Scott Lowe)5 and lung adenocarcinoma (Charles Swanton)6. The role of genetic-environmental interactions has not been studied in detail in GBM, but links to injury are now being uncovered7,8. The finding by Christopher Walsh that the normal human brain contains GBM-associated mutations raises questions about if and how these dormant cells could wake up and give rise to brain cancer9. Could non-genetic events, such as tissue injury or inflammation, push mutant cells into forming a cancer? Our study was also inspired by lineage tracing studies that we had performed in mouse medulloblastoma10,11, and in work by Leonard Zon who demonstrated emergence of developmentally more primitive cells without additional genetic change driving melanoma in a zebrafish model 12.
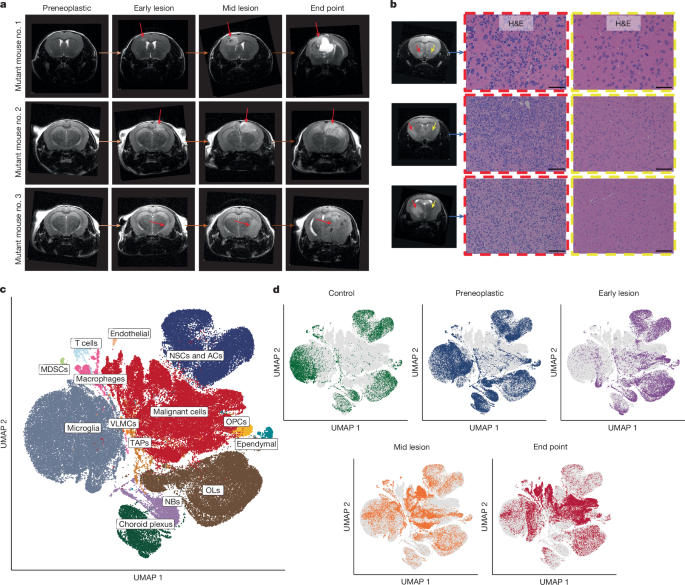
To study the effect of injury-regeneration on GBM formation, we turned to mouse models. In the human brain, there is no possibility to sample preneoplastic or early disease, or to easily generate faithful models from postnatal brain tissues. We turned to a native model where we could study the interactions between an initiated neural cell type and the microenvironment. We utilized a prenatal and postnatal conditional deletion of Trp53 or Trp53/Pten in neural precursors, using Nestin-Cre and Sox2CreER mice, where the initiated cells and their progeny were marked by a tdTomato fluorescent reporter. These deletions represent a large fraction of GBM patients and would be expected to cause mostly disrupted neural lineage function. By studying these mice with serial MRI, it gave us an opportunity to sample reporter-positive and reporter-negative cells representing both malignant and nonmalignant cell states respectively, at all stages of disease, for analysis using single cell approaches (RNA-sequencing and ATAC-sequencing). Importantly, we had recently published a study that defined the cellular states of the neural precursor pool in mice from the embryonic stage to old age, providing us with a map of the precursor cell states and microenvironmental cell types that exist in the brain in steady state conditions13. We were then primed to identify how the transcriptional states of the neural precursors and their progeny change over time during induction and progression of GBM or other environmental perturbation.
By sampling premalignant and malignant precursors throughout the tumourigenic process, we developed a picture of GBM-genesis that revealed some striking findings. First, we found that lesions in the early stages of GBM-genesis, even before there were lesions visible on an MRI, contained cells in the malignant lineage that had acquired expression of a neural crest transcriptional program. Neural crest cells have been recently identified in GBM by others14. Second, a nonmalignant (reporter-negative) population at this very early stage contained cells identifiable by their transcriptional states as disease associated microglia and oligodendrocytes, suggesting that at the earliest stages of the disease, the premalignant neural precursors could be remodeling the gene expression programs of a nonmalignant population. Third, by cell-cell communication analysis, it appears that the neural crest-like mutant population, which shows tremendously open chromatin by ATAC-seq, express ligands that match the expression of receptors on the nonmalignant cells. These findings suggested to us that in the context of neural precursor transformation, one of the early events is the acquisition of a neural crest transcriptional program in the mutant cells. This result was especially interesting because of the known tremendous differentiation and classical brain-organizing potential of normal neural crest cells.
By the time GBM tumours in mice are large, the neural crest-like population is more obscure, being a lower fraction of the cell types compared to early tumours. These cell types, consistent with our previous studies, are highly quiescent. The endpoint tumours become dominated by other neural precursor types that are highly proliferative. By computational analysis, these cells appear to be present in the majority of clones, even at endpoint, and appear to be at the apex of the cellular heterogeneity seen in the tumour.
In parallel to these studies, we wondered how a brain tissue injury affects the transcriptional states of normal neural precursors. We found that a wound that disrupts tissue architecture in the normal mouse brain leads to the appearance of a cell type that expresses neural crest programs, but transiently. The transcriptional programs in these wounded precursors closely resemble that of the early neural crest precursors in the experimental GBMs. These results suggest that the tumour neural-crest like cell types may reflect an injury-like process occurring in early GBM-genesis.
These findings provide fresh insight into gliomagenesis, pointing to the acquisition of a cell state in early tumours highly similar to the states seen after a brain wound. Although much more work is needed, these neural crest-like precursor cells may orchestrate the entire tumourigenic process, by activating surrounding normal cell types, and also by potentially expressing the ligands that act on downstream progeny that seem to acquire the expression of the receptors for neural crest-like cell expressed ligands. This study should bring new ideas to the forefront about the causes of GBM and should suggest new treatments, targeting the roots of GBM growth and the microenvironment that could support tumourigenesis. We believe it also has implications for GBM interception and prevention, by attenuating injury mechanisms at early disease stages. One concept of what may be happening can be seen in the accompanying figure.
We would like to thank our close collaborators who co-authored this study with us: Lincoln Stein and Quang Trinh from the Ontario Institute of Cancer Research, and John Marioni, Kui Hua, and Benjamin Simons from the University of Cambridge, UK.

1 Singh, S. K. et al. Identification of human brain tumour initiating cells. Nature 432, 396-401, doi:10.1038/nature03128 (2004).
2 Lan, X. et al. Fate mapping of human glioblastoma reveals an invariant stem cell hierarchy. Nature 549, 227-232, doi:10.1038/nature23666 (2017).
3 Richards, L. M. et al. Gradient of Developmental and Injury Response transcriptional states defines functional vulnerabilities underpinning glioblastoma heterogeneity. Nat Cancer 2, 157-173, doi:10.1038/s43018-020-00154-9 (2021).
4 Guilhamon, P. et al. Single-cell chromatin accessibility profiling of glioblastoma identifies an invasive cancer stem cell population associated with lower survival. Elife 10, doi:10.7554/eLife.64090 (2021).
5 Alonso-Curbelo, D. et al. A gene-environment-induced epigenetic program initiates tumorigenesis. Nature 590, 642-648, doi:10.1038/s41586-020-03147-x (2021).
6 Hill, W. et al. Lung adenocarcinoma promotion by air pollutants. Nature 616, 159-167, doi:10.1038/s41586-023-05874-3 (2023).
7 Brooks, L. J. et al. The white matter is a pro-differentiative niche for glioblastoma. Nat Commun 12, 2184, doi:10.1038/s41467-021-22225-w (2021).
8 Brooks, L. J., Simpson Ragdale, H., Hill, C. S., Clements, M. & Parrinello, S. Injury programs shape glioblastoma. Trends in neurosciences 45, 865-876, doi:10.1016/j.tins.2022.08.006 (2022).
9 Ganz, J. et al. Rates and Patterns of Clonal Oncogenic Mutations in the Normal Human Brain. Cancer Discov 12, 172-185, doi:10.1158/2159-8290.CD-21-0245 (2022).
10 Vanner, R. J. et al. Quiescent sox2(+) cells drive hierarchical growth and relapse in sonic hedgehog subgroup medulloblastoma. Cancer Cell 26, 33-47, doi:10.1016/j.ccr.2014.05.005 (2014).
11 Selvadurai, H. J. et al. Medulloblastoma Arises from the Persistence of a Rare and Transient Sox2(+) Granule Neuron Precursor. Cell Rep 31, 107511, doi:10.1016/j.celrep.2020.03.075 (2020).
12 Kaufman, C. K. et al. A zebrafish melanoma model reveals emergence of neural crest identity during melanoma initiation. Science 351, aad2197, doi:10.1126/science.aad2197 (2016).
13 Hamed, A. A. et al. A brain precursor atlas reveals the acquisition of developmental-like states in adult cerebral tumours. Nat Commun 13, 4178, doi:10.1038/s41467-022-31408-y (2022).
14 Hu, Y. et al. Neural network learning defines glioblastoma features to be of neural crest perivascular or radial glia lineages. Sci Adv 8, eabm6340, doi:10.1126/sciadv.abm6340 (2022).
Follow the Topic
-
Nature
A weekly international journal publishing the finest peer-reviewed research in all fields of science and technology on the basis of its originality, importance, interdisciplinary interest, timeliness, accessibility, elegance and surprising conclusions.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in