Plants that live longer mast stronger
Published in Ecology & Evolution and Plant Science
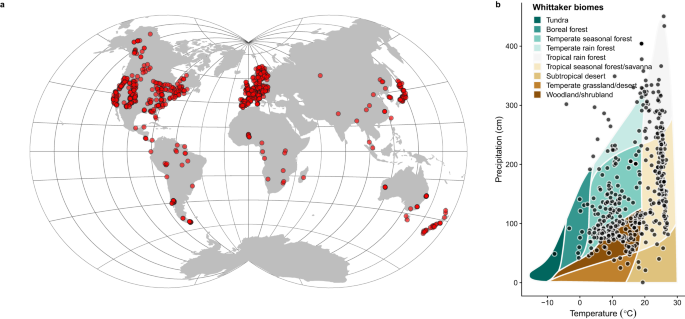
Many plants do not produce seed crops regularly each year. Instead, these plants forgo reproduction for a year or more to produce large crops at irregular intervals, switching between years of abundant seed crops (known as “mast years”) and years with low seed production; this is what we call mast seeding or masting. Intriguingly, these mast years occur simultaneously in plants living alongside each other. That creates boom and bust dynamics in food webs, as seed consumers either have few seeds to feed on and starve, or so many seeds that they are overwhelmed, and some seeds escape being eaten.
These starvation and satiation cycles underpin the fitness benefit that selects for masting. Plants cannot run or hide from their seed predators – instead, masting allows them to escape predation despite standing still. The other benefit of masting is pollination efficiency. Concentrating flowering effort into large events allows plants to saturate the air with pollen, increasing the proportion of flowers that are successfully pollinated.
.png)
But there are costs to masting, and the major one is believed to be skipped reproductive opportunities. In years when seeds are not produced, plants bear two types of related risks. First, a suitable gap (like an opening in the forest canopy) might have just appeared, and there is no seed to colonize that ephemeral opportunity. Other plants, perhaps of species that produce seeds every year, will seize the opportunity. Over time, missing these opportunities could eradicate the masting species from otherwise suitable locations. The second associated risk is dying before reproduction happens, wasting the saved resources. Crucially, both these costs are lower if you live longer. For a plant that lives hundreds of years, the chances of reproducing at the same time as a gap appearing is higher, because you reproduce more times across your lifetime. Moreover, as the probability of surviving might be 99% per year in long-lived species, the risk of dying before the mast year is negligible. However, while it has proven relatively easy to test the benefit of masting -- for example by measuring the proportion of predated seeds or pollinated flowers as a function of crop size, the task is more difficult when it comes to measuring the costs. One way would be to relate the strength of masting (how much reproduction varies across years) to survival rates of plant species. We would expect stronger masting in species that live longer. But that’s not easy. Data on seed production for a large enough sample of species was not available until very recently. Secondly, data on survival rates are similarly difficult to get at such a scale.
First, we solved the seed production data issue. In 2018, we started drawing together published data from the literature and our own datasets. We developed the largest published masting database as part of the MAST-NET project. This dataset – MASTREE+ – contains over 70,000 records of plant reproduction from almost 1000 species. In the paper now published in Nature Communications, we have merged this masting data with species functional traits - indicators of life history strategies.
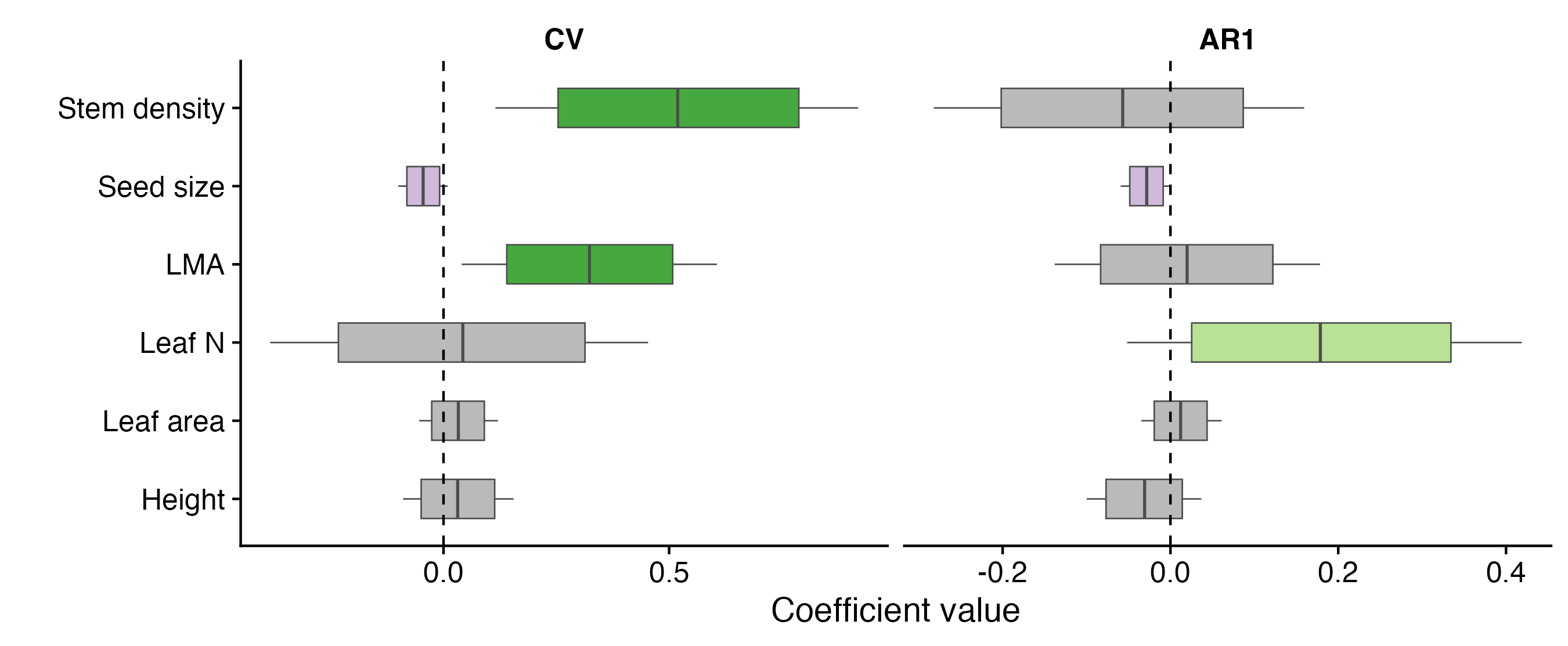
The main functional trait of interest was wood density (“stem tissue density” in the article, as not only woody plants were included). Research has repeatedly demonstrated that high wood density provides mechanical strength and reduces mortality, but limits growth rates, which distinguishes strategies reliant on stress persistence from those based on rapid utilization of ephemeral opportunities. We thus expected stronger masting in species with high wood density, as lower mortality rates due to stronger stress resistance should buffer against the missed reproductive opportunities that are the main costs of masting.
Linking these two - masting strength and functional traits - is, however, complicated by their relationships with climate and phylogeny. Species may mast strongly and have high wood density not because wood density enabled the evolution of masting, but because particular climates happen to select for both independently. Similarly, closely related species may share strong masting and high wood density because their ancestors happened to have both of these traits. The problem of such an indirect relationship is apparent as there are well-known geographical gradients in masting and wood density. In the tropics, masting is weaker and wood density is high. We needed a tool to separate these confounding factors.
Generalized joint attribute modeling (GJAM) comes to the rescue, allowing the decomposition of direct and indirect relationships, driven by either climate or phylogeny. GJAM models indicated that while interannual variation in seed production is highest in temperate and boreal zones, controlling for environment and phylogeny reveals that masting is more frequent in species that invest in traits that promote long lifespan. Our modeling exposed masting-functional trait relationships that would otherwise remain hidden, and provides large-scale evidence that the costs of delayed reproduction play a significant role in the evolution of variable reproduction in plants.
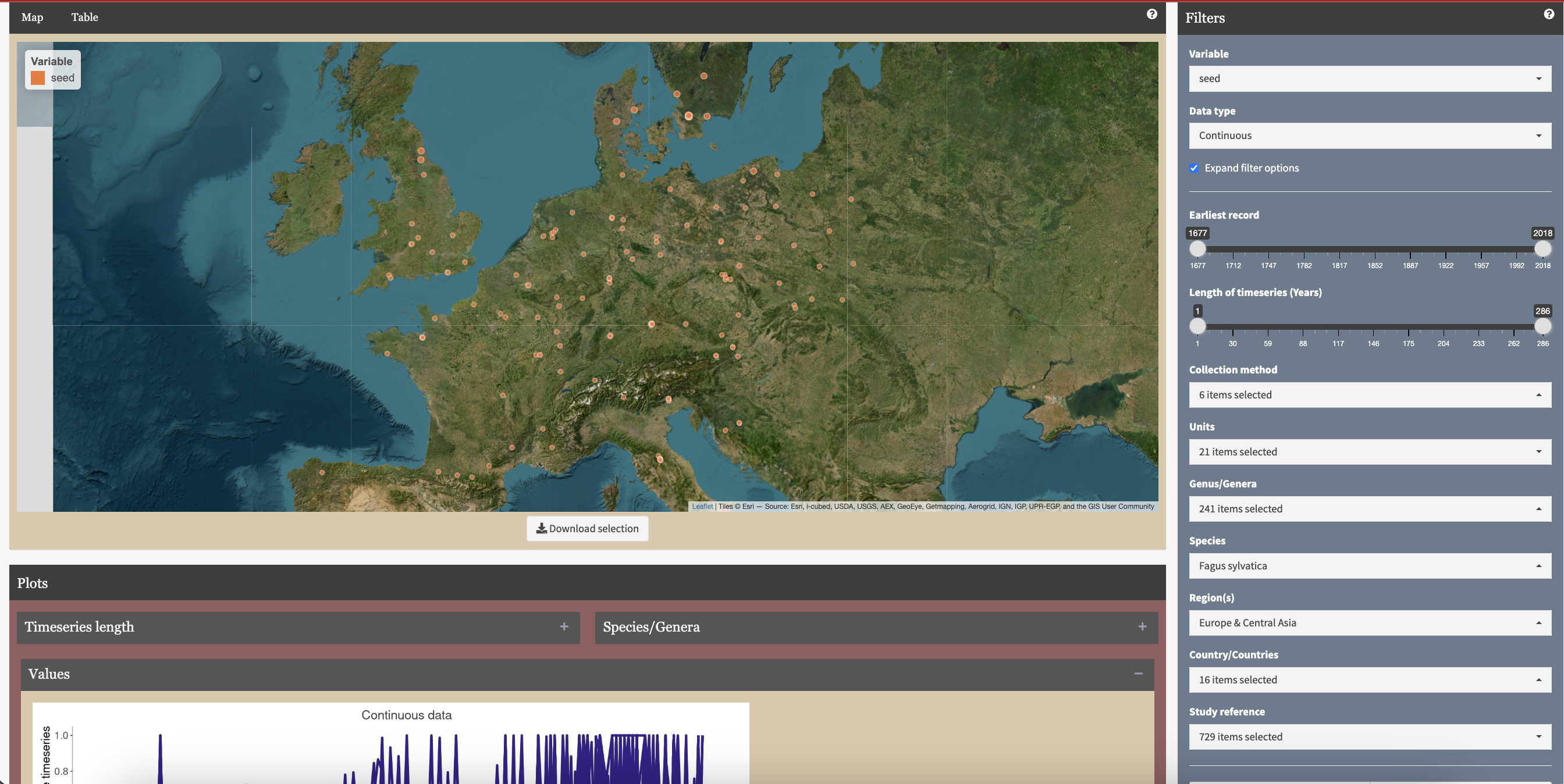
The MASTREE+ database is open access, and all the data can be easily explored and accessed with the following application (Illustration 3). The large coverage of species and the high replication of some of them, allows new questions to be explored. In addition to the study published in Nature Communications, MASTREE+ has been used to demonstrate the potential for mast forecasting and we also used the dataset to discover why masting in some species is synchronized at distances reaching 1500 km, while synchronization is more local in others. We welcome anyone interested in plant reproductive ecology to explore the data!
Other related references :
Hacket-Pain, A., Foest, J. J., Pearse, I. S., LaMontagne, J. M., Koenig, W. D., Vacchiano, G., Bogdziewicz, M., Caignard, T., Celebias, P., van Dormolen, J., Fernández-Martínez, M., Moris, J. V., Palaghianu, C., Pesendorfer, M., Satake, A., Schermer, E., Tanentzap, A. J., Thomas, P. A., Vecchio, D., … Ascoli, D. (2022). MASTREE+: Time-series of plant reproductive effort from six continents. Global Change Biology, 28(9), 3066–3082. https://doi.org/10.1111/gcb.16130
Bogdziewicz, M., Journé, V., Hacket-Pain, A., & Szymkowiak, J. (2023). Mechanisms driving interspecific variation in regional synchrony of trees reproduction. Ecology Letters, 26(5), 754–764. https://doi.org/10.1111/ele.14187
Journé, V., Hacket-Pain, A., Oberklammer, I., Pesendorfer, M. B., & Bogdziewicz, M. (2023). Forecasting seed production in perennial plants: identifying challenges and charting a path forward. New Phytologist, 239(2), 466–476. https://doi.org/10.1111/nph.18957
I am a postdoctorate, interested in ecology and forests. I like patterns in ecology (e.g. seed production, species distribution) and general theory in ecology tested with observational data. For now, I am working more on seed production in perennial plants, and I am trying to understand why plants are reproducing or not - 'to reproduce, or not to reproduce, that is the question' Masting, Act 3, Scene 1. I like also using different statistical tools and methods
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in