Publication investigating the dual substrate specificty of the PRDM9 protein lysine methyltransferase
Published in Cell & Molecular Biology
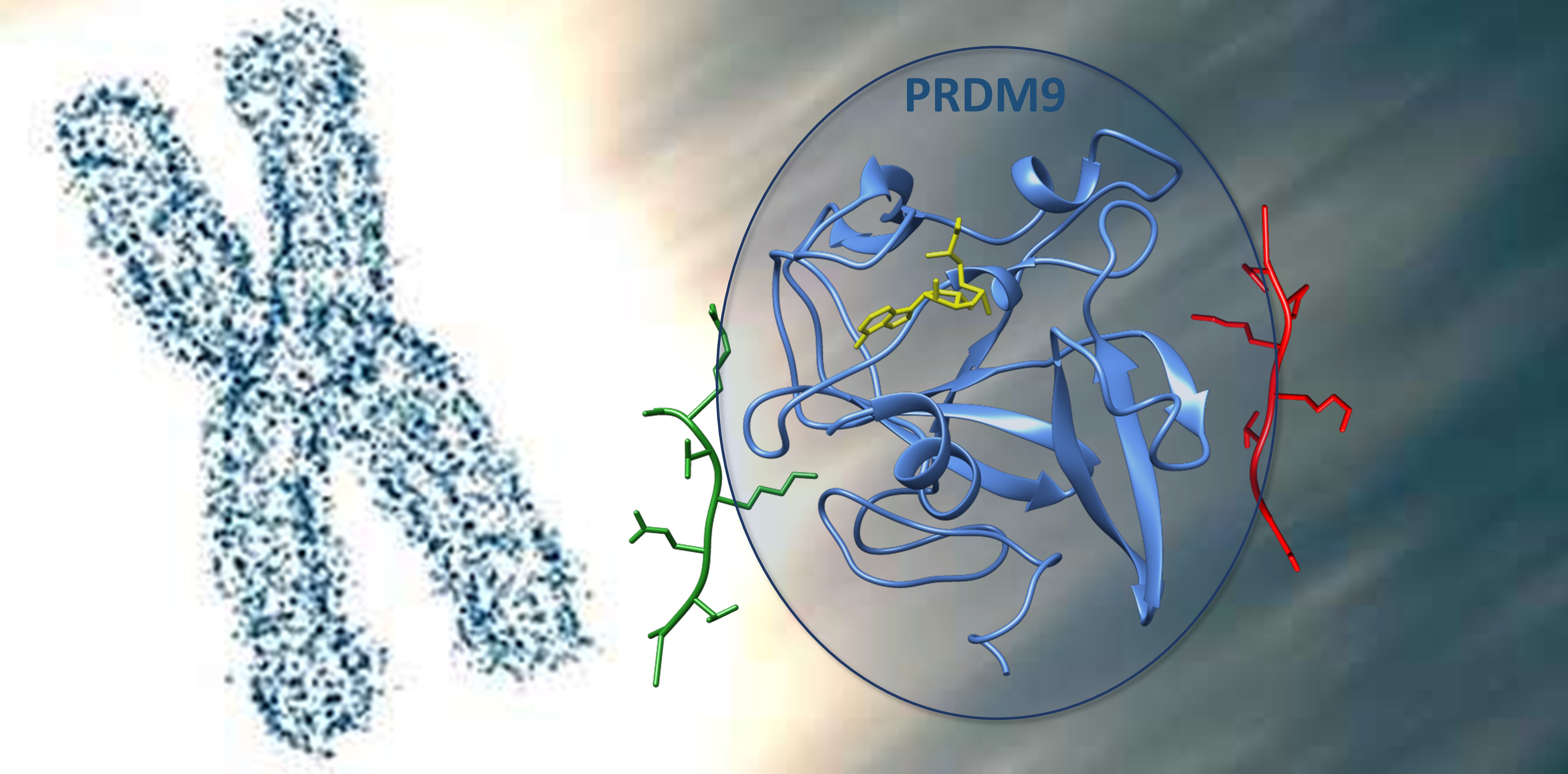
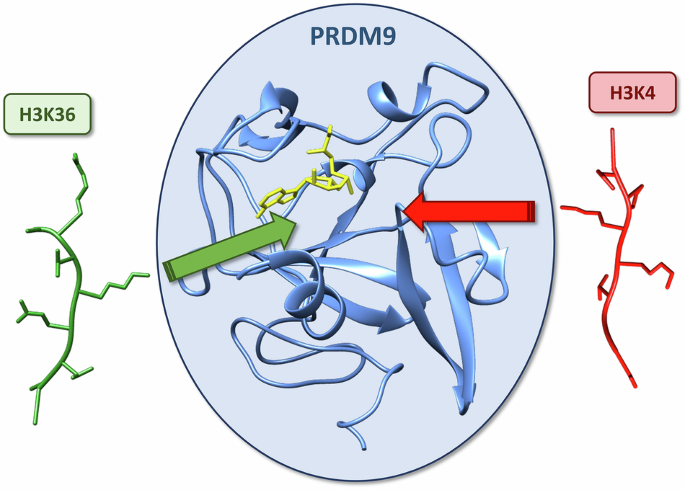
Summary
The PRDM9 protein lysine methyltransferase is essential in meiotic recombination where it trimethylates H3K4 and H3K36 in chromatin. However, it is not known how this enzyme can specifically methylate these two substrates despite their dissimilar amino acid sequences. By combining biochemical and modelling techniques, we show here that this exceptional dual substrate specificity is based on a bipartite peptide recognition cleft, comprising one peripheral binding site specific for the H3K4 peptide but tolerating H3K36, and one central binding site with the opposite preferences. The N-terminal part of the H3K4 peptide and R2 are bound by a network of PRDM9 residues in a bent conformation that sterically excludes the continuation of the peptide chain. The H3K36 peptide contains G33 and G34 in this part allowing the peptide chain to follow the binding path of the R2 side chain in the H3K4 complex. Because of this, a continuous peptide can be bound in the H3K36 binding mode. Conversely, in the central part of the substrate binding pocket, hydrophobic residues as those found in H3K36 are highly preferred, enabling an accurate readout and efficient methylation of the H3K36 sequence. Still, the residues found in H3K4 in this region are tolerated which in combination with the strong recognition of the N-terminal part of H3K4 in the peripheral region of the binding site allows to specifically methylate the H3K4 substrate as well. Based on our data, site-directed mutagenesis of residues involved in PRDM9-peptide contacts allowed us to modulate the K4/K36 preferences strongly. In our protein engineering experiments, PRDM9 mutants were identified with elevated preference for H3K4 (H3K4 >> H3K36), with lost preference (H3K4 ≈ H3K36), and even with inverted preference (H3K36 > H3K4). One additional mutant showed very low activity I339Q mutant, but in this case misfolding of the purified protein cannot be excluded. These findings document that the remarkable substrate recognition by PRDM9 with dual specificity can be tuned towards the preference for one or the other peptide.
Detailed description
Histone lysine methylation is a key post-translational modification that regulates gene expression. Two such modifications—H3K4me3 and H3K36me3—are typically catalyzed by distinct lysine methyltransferases (PKMTs). However, PRDM9 uniquely catalyzes the trimethylation of both H3K4 and H3K36 at meiotic recombination hotspots, despite the dissimilarity of their amino acid sequences. The molecular basis for this dual specificity has remained unclear. The present study aimed to decipher the structural and biochemical basis of this dual specificity through a combination of peptide methylation assays, site-directed mutagenesis, and molecular dynamics (MD) simulations.
Purified His-tagged PRDM9 was shown to methylate both H3K4 and H3K36 peptides, including their mono- and dimethylated forms. No methylation occurred on lysine-to-alanine mutants, confirming sequence-specific methylation of K4 and K36. Using SPOT arrays, each position of the H3K4 (residues 1–15) and H3K36 (29–43) peptides was substituted by all proteinogenic amino acids (except Trp and Cys). The methylation levels of each variant peptide revealed distinct and position-specific preferences.
- H3K4 context: Strong preferences at -3 (Ala), -2 (Arg), and +2 (Val) sites suggest tight, selective interactions.
- H3K36 context: Preferences are less strict at the N-terminus, but strong at -1 (Val), +1 (Lys), and +2 (Pro), suggesting these sites are critical for H3K36 recognition.
Overall, PRDM9 appears optimized to accommodate both sequences through a dual specificity in a bipartite binding cleft to explain the dual substrate specificity. The N-terminal region of the H3K4 peptide is bound tightly at A1 and R2, which forces a conformation that prevents peptide extension. In contrast, the H3K36 peptide carries glycine residues (G33 and G34) that allow a more extended conformation. The central region of the peptides (-1 to +2) shows stronger interactions in case of H3K36 peptide residues (e.g., V35, K37, P38), which are hydrophobic or positively charged. H3K4 peptide residues are tolerated but less preferred in this region. This arrangement enables PRDM9 to methylate both substrates with distinct interaction modes.
MD simulations of PRDM9 in complex with either H3K4 or H3K36 peptides revealed differential peptide binding conformations:
- H3K4: R2 fits into a narrow channel and interacts with E360 and Y361. This causes the N-terminus to adopt a bent, terminal conformation.
- H3K36: G33 and G34 occupy the R2 channel, permitting the peptide to extend along the PRDM9 surface.
At the -1 and +1 positions, hydrophobic interactions (e.g., with A287, L294) and electrostatic contacts (e.g., K37 with E360) further differentiate the two binding modes. TS-like conformations, essential for methylation catalysis, were observed ~4× more frequently in H3K4 simulations, consistent with the ~5× higher catalytic rate for H3K4 in biochemical assays.
Seven key residues of PRDM9 involved in substrate contact were mutated:
- A287S: Diminished overall activity, especially with H3K36, thus increasing preference for H3K4.
- Y361A & E364A: Both mutations reduced H3K4 activity more than H3K36 activity. The E364R mutant abolished H3K4 activity, fully inverting specificity to favor H3K36.
- E360 variants: E360K and E360P suppressed H3K36 methylation, increasing H3K4 preference. E360D enhanced H3K36 activity, reducing the H3K4/H3K36 ratio.
- L294S: Increased H3K36 activity while preserving H3K4 activity.
- I339Q: Severely reduced activity on both peptides, perhaps due to misfolding.
- F333A/Y/W: All these mutants significantly increased H3K36 methylation without compromising H3K4 methylation, highlighting F333’s role in steric constraints at the +2 position.
These mutations demonstrate that altering single residues can profoundly influence substrate preference, offering insight into possible evolutionary paths of PKMT diversification. Among the numerous human SET- or PRDM-domain containing PKMTs many display distinct substrate specificities. The PRDM9 mutants created in this study illustrate how subtle sequence changes within the SET domain can modulate substrate preference. This supports a model where PKMTs evolved new specificities via few but critical mutations, enabling adaptation to diverse functions. This study thus advances our mechanistic understanding of dual specificity in PKMTs and provides a foundation for reengineering methyltransferase specificity, with broad implications in epigenetic research and enzyme design.
I am Professor of Biochemistry at University of Stuttgart, where I am heading the Institute of Biochemistry. My group investigates the biochemistry and enzymology of DNA and protein methyltransferases. In the field of Molecular Epigenetics, my group studies DNA and chromatin modifications and the specificity of reading domains. Built on a long-standing expertise in protein design of DNA interacting enzymes, we also develop and apply systems for epigenome editing in eukaryotic cells.
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026
DNA repair and human disease
Publishing Model: Hybrid
Deadline: Oct 31, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in