T cells and big bites are all B cells need.
Published in Healthcare & Nursing
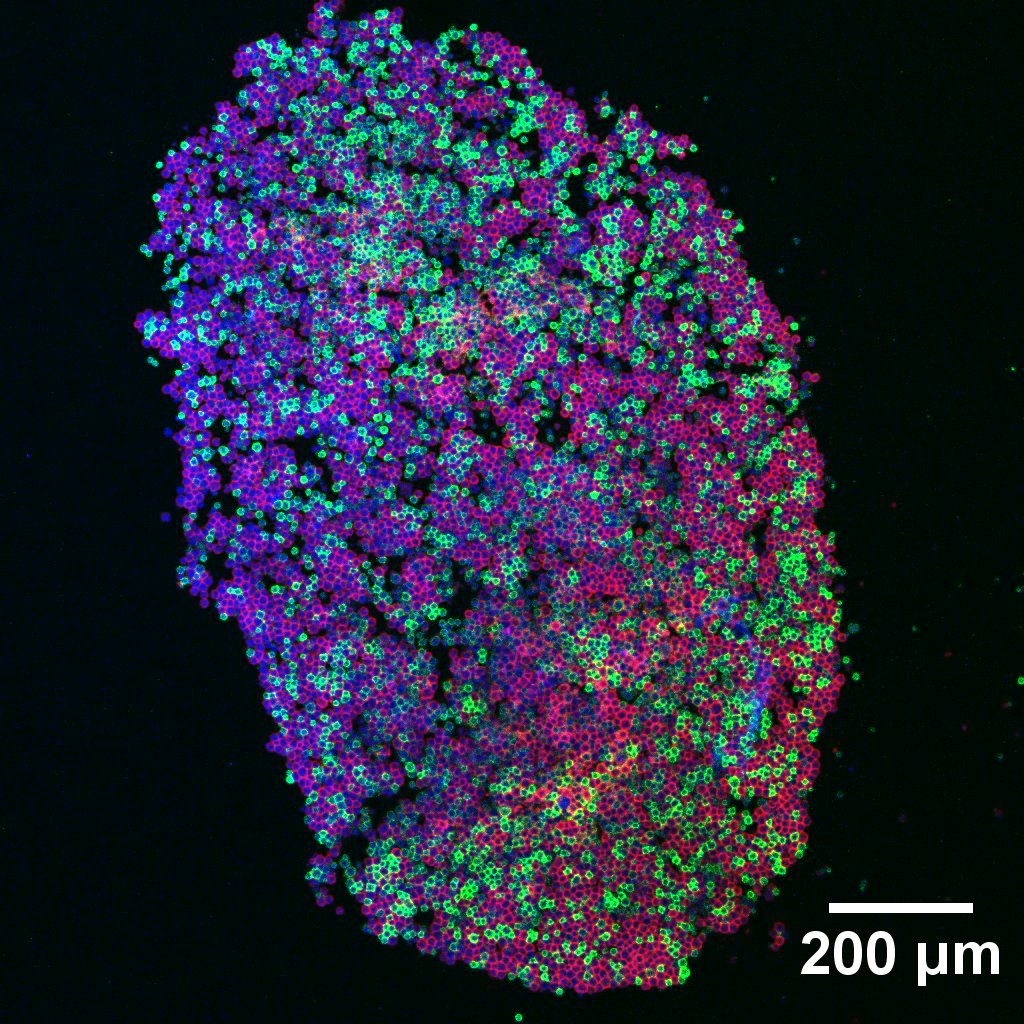
The production of high-affinity antibodies capable of targeting and eliminating foreign pathogens characterizes the humoral arm of our adaptive immune system. To induce this protective response against pathogens, B lymphocytes must undergo a dynamic process called the germinal center (GC) reaction, which takes place in the B cell follicles of the secondary lymphoid organs. There, a Darwinian process based on mutation and selection favors the survival of those B cells that carry B cell receptors (BCRs) with higher affinity for the antigen. To be selected, B cells must receive two important signals, the first from the BCR through recognition of the cognate antigen, and the second from cognate CD4 T cells differentiated into T follicular helper cells (Tfh). B cells that receive the positively selecting signals from T cells undergo further rounds of somatic hypermutation on their B cell receptor genes to increase their affinity for the antigen. The GC output generates effector memory and plasma cells, which tend to emerge at slightly different time points in the response and carry BCRs of different affinities1.
Despite the importance of the GC, its complexity and dynamics have hindered a deeper analysis of the process, leaving many questions unanswered, such as which molecular signals control the multiple differentiation steps. The use of in vitro systems to mimic in vivo processes has been extensively developed in different scientific fields to study complex processes with simpler experiments. As a result, several groups have reported systems that mimic GCs in vitro 2,3. However, some of the most relevant steps of the GC process could not be recapitulated with these systems.
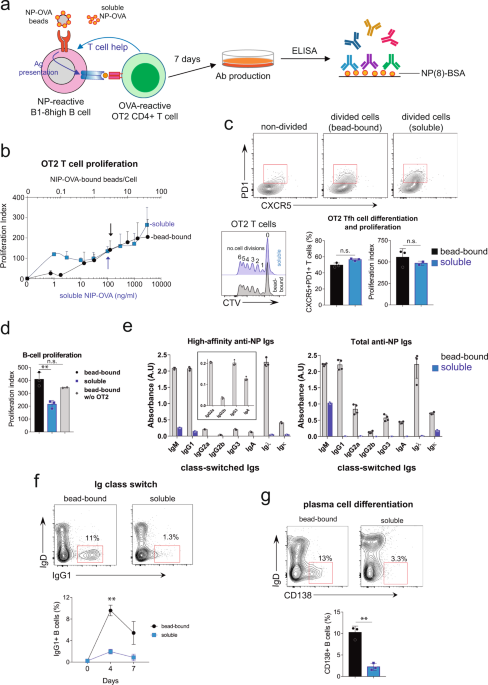
In our publication, we provide a new methodology to reconstitute GCs in vitro (Martinez-Riano et al, 2023). This new approach is based on phagocytic delivery of antigen to B cells and co-culture with cognate CD4 T cells, providing the two essential signals for B cells to differentiate into GC cells. In this case, phagocytic antigens generate a stronger and more persistent BCR signal than soluble antigens, resulting in enhanced in vitro GC differentiation. The novelty of our method is that phagocytic antigen uptake facilitates affinity selection within the cultured B cells, since those cells bearing higher affinity BCRs have a cytoskeletal-remodeling advantage to engulf large antigens compared to cells bearing lower affinity antigens. In our co-culture conditions, cognate naive CD4 T cells differentiate into Tfh cells and support the differentiation and survival of the GC B cells. Under these conditions, we see that differentiated GC B cells further differentiate into class-switching antibody-producing plasma cells that secrete high-affinity antibodies against the antigen provided in culture.
This new method to reconstitute the GC in vitro aims to facilitate the study of this complicated process by allowing high-throughput screens to be performed and specific molecular mechanisms to be analyzed in a time-controlled manner. As an example, we were particularly interested in studying the importance of T-B contacts within the GC. The rationale was that soluble signals secreted by Tfhs upon stimulation by cognate high-affinity B cells could also modify bystander unrelated B cells, or even autoreactive B cells located nearby. We therefore specifically analyzed the relevance of T cell help to B cells in a direct or bystander manner in the co-culture. Surprisingly, while cognate B cells differentiate into plasma cells, bystander B cells acquire a memory B cell phenotype, suggesting that memory B cell differentiation may require only some of the signals provided by T cells.
In conclusion, we believe that our work provides a new tool for immunologists to study the GC response in a more accessible manner and supports the ability to generate high affinity antibodies in vitro against antigens of interest for vaccines. The key for GC B cells to succeed in a Petri dish is to eat large antigens and hug tightly to T cells.
References
- Victora, G. D. & Nussenzweig, M. C. Germinal centers. Annu Rev Immunol. 30:429-57., 10.1146/annurev-immunol-020711-075032. Epub 2012 Jan 3. (2012).
- Nojima, T. et al. In-vitro derived germinal centre B cells differentially generate memory B or plasma cells in vivo. Nat Commun. 2:465., 10.1038/ncomms1475. (2011).
- Ochiai, K. et al. Protocol for in vitro BCR-mediated plasma cell differentiation and purification of chromatin-associated proteins. STAR Protocols 2, 100633 (2021).
My most important scientific contributions are framed in two areas of research: immunology and cancer. Among my various contributions in immunology, perhaps the most outstanding from an intellectual and subsequently translational point of view, has been the demonstration that the receptor for the antigen of T lymphocytes (TCR) undergoes conformational changes after the binding of antigens and agonistic antibodies (Cell, 2002). This discovery broke the existing dogma that the TCR did not undergo conformational changes, but rather signal transmission occurred through mere mechanisms of aggregation and/or molecular exclusion. This discovery has revolutionized the view of the TCR, perhaps the most important molecule in the functioning of the adaptive immune response. In fact, and just as an example, the TCR enables vaccines to work or for so-called "checkpoint inhibitors" to have anticancer activity. It is also part of the development of cancer therapy known as "CAR-T". We based the demonstration of the existence of conformational changes in the TCR on the observation that, as a consequence of antigen stimulation, the TCR exposes a sequence that recruits the adaptor protein Nck. This occurs through an SH3 domain of Nck that has a unique "pocket". This offered the opportunity to design molecules that interfere with the TCR-Nck binding. Following this idea, we designed, through an "in silico" procedure, a low molecular weight inhibitor to fit into that pocket and prevent the recruitment of Nck to the TCR, and therefore signal transmission. Such inhibitor, which we named AX024, proved to be effective through oral administration in various models of autoimmune diseases: psoriasis, Crohn's disease/ulcerative colitis, asthma, and multiple sclerosis (Sci. Transl. Med, 2016), all caused to a large extent by uncontrolled activation of T lymphocytes. This inhibitor and its entire family of derivatives were patented and led to the creation of the company Artax Biopharma Inc based in Boston MA. I am a founding member of Artax, I was its CSO and now I am a member of its Scientific Advisory Board. Artax has improved the initial compound with the lead AX158. This latter compound has completed all regulatory preclinical phases and Phase I. It is currently in Phase II for psoriasis treatment. If the drug meets expectations, it will be the first immunomodulatory drug, not immunosuppressive, acting on the TCR and with a possibility of use in practically all autoimmune diseases. The drug is administered orally, with only a daily dose and unaffected by food intake or not. In conclusion, it is a "First-in-class" drug for many reasons: it modulates TCR signaling and acts by inhibiting a protein-protein interaction, not an enzymatic activity, which distinguishes it from most pharmacological compounds currently in use. Our interest in characterizing TCR signaling molecules led us to identify not only Nck but also R-RAS2; obtaining novel and impactful results in cancer. R-RAS2 is a GTPase similar to the oncogene K-RAS, which, however, does not undergo activating mutations in practically any type of cancer. Perhaps for this reason, interest in R-RAS2 declined despite being discovered in 1990. Contrary to K-RAS and other similar GTPases, R-RAS2 has a high intrinsic activity of guanosine nucleotide exchange. This peculiarity led us to think that elevated levels of R-RAS2 expression, even in the absence of activating mutations, could cause cancer. Our work, based on the generation of a transgenic mouse that overexpresses the wild-type form of R-RAS2, demonstrated that this hypothesis is correct. In fact, we found that all mice develop chronic lymphocytic leukemia (CLL), a B-cell leukemia that is the most common in the West (Mol Cancer, 2022). This allowed us to translate these observations to humans, analyzing a cohort of patients with CLL. This study showed that more than 80% of patients overexpress non-mutated R-RAS2. Additionally, we found genetic markers in non-coding regions of the RRAS2 gene that are linked to this overexpression. Such markers can be used as a prognostic method as they are linked to worse survival. Currently, we are characterizing RRAS2 as the most frequently implicated gene (68%) in the development of breast cancer, especially in forms known as "triple-negative" and those linked to pregnancy. In addition to these two lines of research representing the pillars of my research career, we have been involved in the creation of a diagnostic test for immunity to SARS-CoV-2 that was patented and commercialized by a Spanish company during the recent pandemic (EMBO Mol Med, 2021). The method on which this test is based can be used for the evaluation of humoral response to any pathogen. In summary, I believe that my trajectory shows how research aimed at understanding basic mechanisms can be translated into clinical use. One of the products of this translation, AX158, has reached the Clinic, and another product, genetic markers of RRAS2, is in the process of materializing as a prognostic marker, in addition to pointing to R-RAS2 as a target for future therapies.
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Mechanistic insights into human host and microbiome interactions
Publishing Model: Open Access
Deadline: May 31, 2026
Advances in neurodegenerative diseases
Publishing Model: Hybrid
Deadline: Jun 30, 2026



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in