Salmonella enterica is probably the best-known bacterium that causes food-poisoning. In fact a large number of cases arise from the consumption of contaminated meat, as well as eggs and sometimes fresh vegetables and fruit. While many pathogenic varieties of Salmonella enterica exist, they don't all look the same! Some of them have evolved versatile strategies to trick their hosts’ immune defences and escape from the gut, causing invasive disease. These types of Salmonella are of most concern as they are responsible for severe disease. In my opinion, Salmonella Typhi is without doubt the most fascinating one. As compared to non-typhoidal Salmonella, it can hide inside the immune cells and hitch a ride to different organs and body sites where it can cause infection. It can also trigger overburdening immune reactions and produce large quantities of toxins. Mostly important, it has adapted strictly to human hosts, making this pathogen particularly difficult to study in other model organisms such as mice.
Once ingested, Salmonella reach the small intestine and go inside many cells including epithelial cells and immune cells such as macrophages and dendritic cells (DCs). DCs are the “sentinels” of the immune system and have the central ability to initiate or silence an immune response, but Salmonella, smartly enough, has evolved mechanisms to manipulate the DC immune response. Defining the differences between molecular interactions of mild and severe variants of Salmonella and host DCs is important in finding ways to prevent or treat these bacterial infections more effectively.
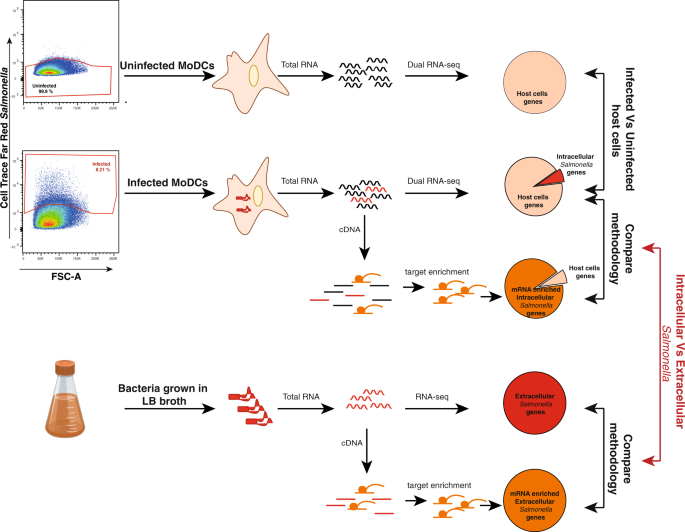
But how can we take a snapshot of the crosstalk between Salmonella and host DCs?
Understanding how bacteria cause disease requires knowledge of which genes are expressed and how they are regulated during infection. Yet, this is not enough, because organisms do not exist isolated from each other, but constantly interact with each other. In our case, invasive bacteria must adapt to the environment of the host cell to survive while the host cell reacts to the invader in an attempt to clear the infection. Fortunately, recent advances in next-generation sequencing over the last couple of years allowed us to apply RNA sequencing to two organisms at the same time (dual RNA-seq), and thus to simultaneously study the gene expression of two interacting species.
Initially we planned to infect human DCs with mild and severe Salmonella and, in a simple way, to sequence all the transcripts present in the infected cells at 6 hours post-infection. As frequently happens in science, we realized that our plan needed to be improved: given the vast excess of human RNA transcripts compared to bacterial we would end up with very little information from Salmonella. The solution was quickly suggested by our computational biologist: why don’t we design a set of probes targeting Salmonella transcriptome that we can use as a bait to fish out bacterial genes? This approach turned out to be a winning strategy: we greatly enhanced the recovery of bacterial sequences with those probes.
The most striking result that emerged from our data was a specific pathway involved in iron acquisition and storage activated uniquely by DCs following infection with S. Typhi. Iron is critical for the biochemistry of cells and is needed equally by host and pathogen, indeed the competition for iron is a key determinant of the outcome of infection. On one side, iron is essential for the growth of intracellular pathogens, while on the other side it is also a major source of reactive species of oxygen and nitrogen, toxic components that destroy bacteria and help the immune system to clear infections.
With this in mind, we wondered how iron might influence the anti-bacterial response of DCs and if the outcome might be different for mild and invasive Salmonella. In a time course experiment, we observed that while DCs supplemented with iron actually fared better with guarding against Salmonella, iron-deprived DCs were indeed more susceptible to the spread of the bacteria. It was exciting to see that these results, however, were only applying to non-typhoidal Salmonella. S. Typhi was growing inside DCs undisturbed by the surrounding iron fluctuation. This led us to suspect that S. Typhi could have a trump card that allows it to survive to iron loaded DC that other Salmonella do not have.
Meticulous research in the intricate web of genes expressed by the bacterium during infection pointed out not one but three possible tricks that S. Typhi might use to deceive the iron-induced killing. S. Typhi displayed increased expression of katG, ygaD, and hmpA, three important genes which contribute to detoxify reactive species of oxygen and nitrogen. Encouraged by a peculiar resistance that S. Typhi manifested in the presence of nitrogen stressor, we decided to construct a S. Typhi mutant strain deficient to express the hmpA gene. It was at this point that we observed what was probably the most exciting result of this project: the mutant failed to survive in DCs loaded with iron, supporting the hypothesis that this gene may play a role in typhoidal resistance mechanisms against iron induced stress.
It felt like we added another piece to the puzzle picturing the mechanisms of Typhoidal virulence.
But isn’t iron also an essential micronutrient for bacteria? Salmonella feeds upon iron by using special molecules called siderophores that bind to the metal and rip it right out of the proteins in which it is enclosed. Siderophores then transport the precious cargo back to the bacteria. Given the high level of iron uptake in infected DCs, we were not surprised at all to see that S. Typhi did not show particularly high expression of genes involved in iron acquisition. Plenty of iron was already available! However, it did show particular induction of the non-ribosomal protein synthetase entF. Since EntF has been shown to confer resistance to H2O2 in Salmonella, we hypothesized that S. Typhi siderophore production may be another stratagem adopted by this bacterium to reduce the oxidative and nitrosative stresses. A simple colorimetric assay to measure Salmonella siderophore production confirmed our hypothesis: S. Typhi maintained siderophore production unperturbed by exposure to oxidative or nitrositive stressor.
Nevertheless, the small intestine is the natural site of Salmonella infection and we couldn’t help but wonder how broadly applicable our in vitro results were. Luckily, our group has long experience working with intestinal tissue and we were able to develop an ex vivoinfection model using human terminal ileo biopsies. In this model, bacteria infect tissue myeloid cells, which are known to have unique phenotypes compared to macrophages or DCs derived in vitro from circulating monocytes. Needless to say, we were thrilled to observe that, similar to DCs, ileum cells infected with S. Typhi expressed higher levels of the iron uptake and storage machinery compared to non-typhoidal species, while limiting iron export.
Our dual RNA-seq data highlighted a dynamic adaptation of iron metabolism in both host and pathogen during infection. Given the involvement of iron in the control of microbial pathogenicity, it appears reasonable to propose that targeting iron may offer therapeutic value.
. Excess of iron can be used by the host to generate antimicrobial mechanisms which improve bacterial clearance (2) Infection with S. Typhi results in increased iron loading and activation of oxidative and nitrosative stress mechanisms (1-4). However, the higher production of detoxifying enzymes, such as HmpA (5), protect the bacteria by the toxic effect of RNI. Enterobactin (entF and entC) are responsible of iron acquisition during stress (5)")
Infection with S. Typhi results in increased iron loading and activation of oxidative and nitrosative stress mechanisms (1-4). However, the higher production of detoxifying enzymes, such as HmpA (5), protect the bacteria by the toxic effect of RNI. Enterobactin (entF and entC) are responsible of iron acquisition during stress (5).
Follow the Topic
-
Communications Biology

An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.
Related Collections
With Collections, you can get published faster and increase your visibility.
From RNA Detection to Molecular Mechanisms
Publishing Model: Open Access
Deadline: May 05, 2026
Signalling Pathways of Innate Immunity
Publishing Model: Hybrid
Deadline: May 31, 2026



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in