Unexpected predominance of Archaea in a cool hyperacidic environment.
Published in Microbiology
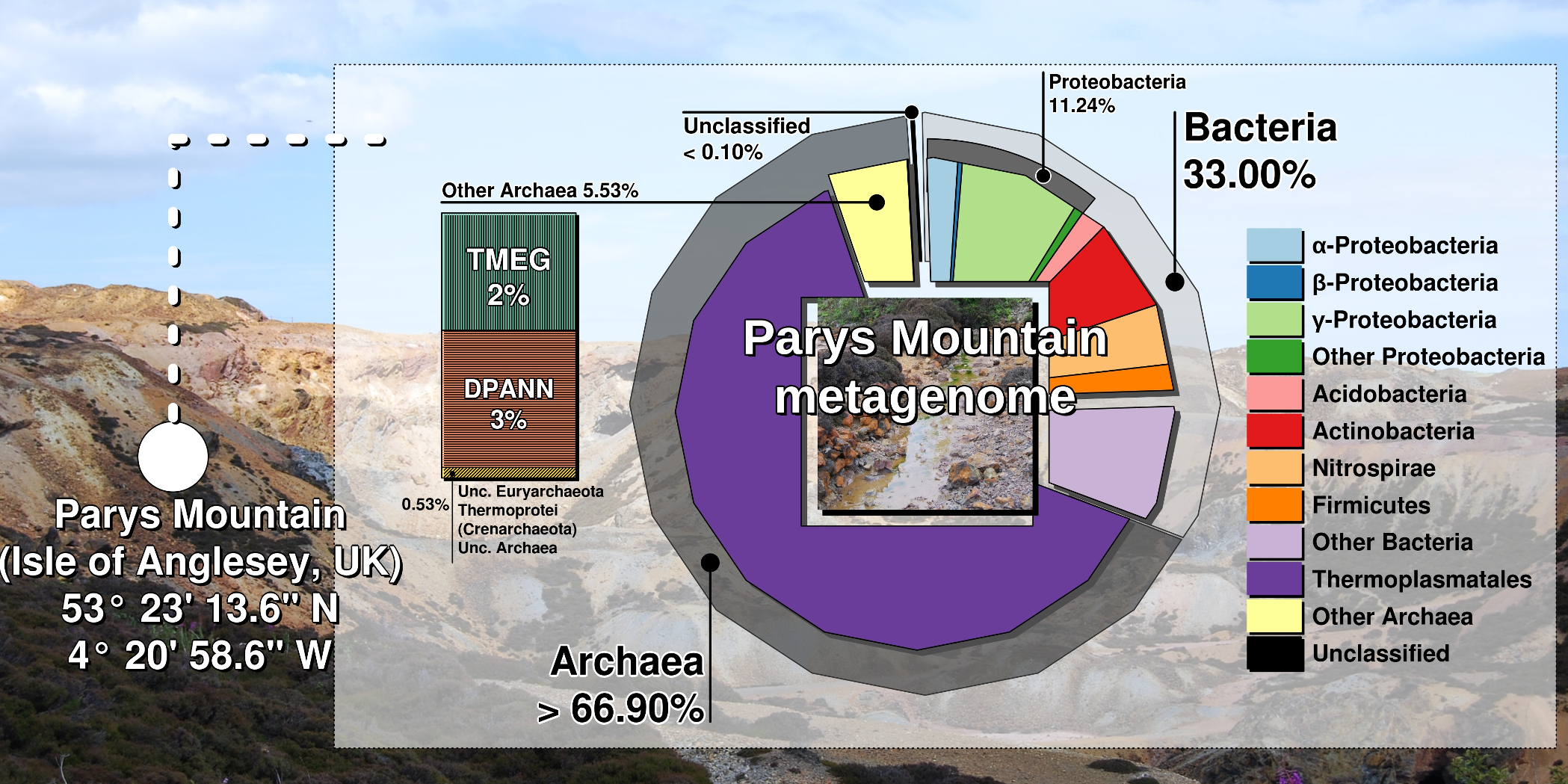

From this very acidic site (pH 1.3-1.6) we have isolated and characterised a new archaeal genus and species, Cuniculiplasma divulgatum from a new family Cuniculiplasmataceae (order Thermoplasmatales). Comparative genomic analysis of Cuniculiplasma strains, which we isolated from Parys Mountain and Iberian pyritic belt site and a metagenomic assembly from a sibling organism from California pointed at a remarkable degree of conservation in their genomes (1,2).
Later, we described the association between C. divulgatum and its ectosymbiont (or ectoparasite) “Candidatus Mancarchaeum acidiphilum” (“Micrarchaeota”) and reported on the analysis and evolutionary patterns of this first ungapped genome in DPANN Superphylum (3). It was quite logical that our next step towards better understanding of microbial diversity of this hyperacidic ecosystem would be the assessment of the entire acidophilic microbiome and spatial distribution of microorganisms inhabiting water and sediment subsystems.
Shotgun metagenomics and SSU rRNA amplicon sequencing of DNA samples revealed that Euryarchaeota accounted for 67 % from the whole community, with the prevalence of Thermoplasmata (58%). Within Thermoplasmata, one particular group, “E-plasma”, without isolated representatives was in a majority (up to 43.5 % of total metagenomic reads) (Fig).
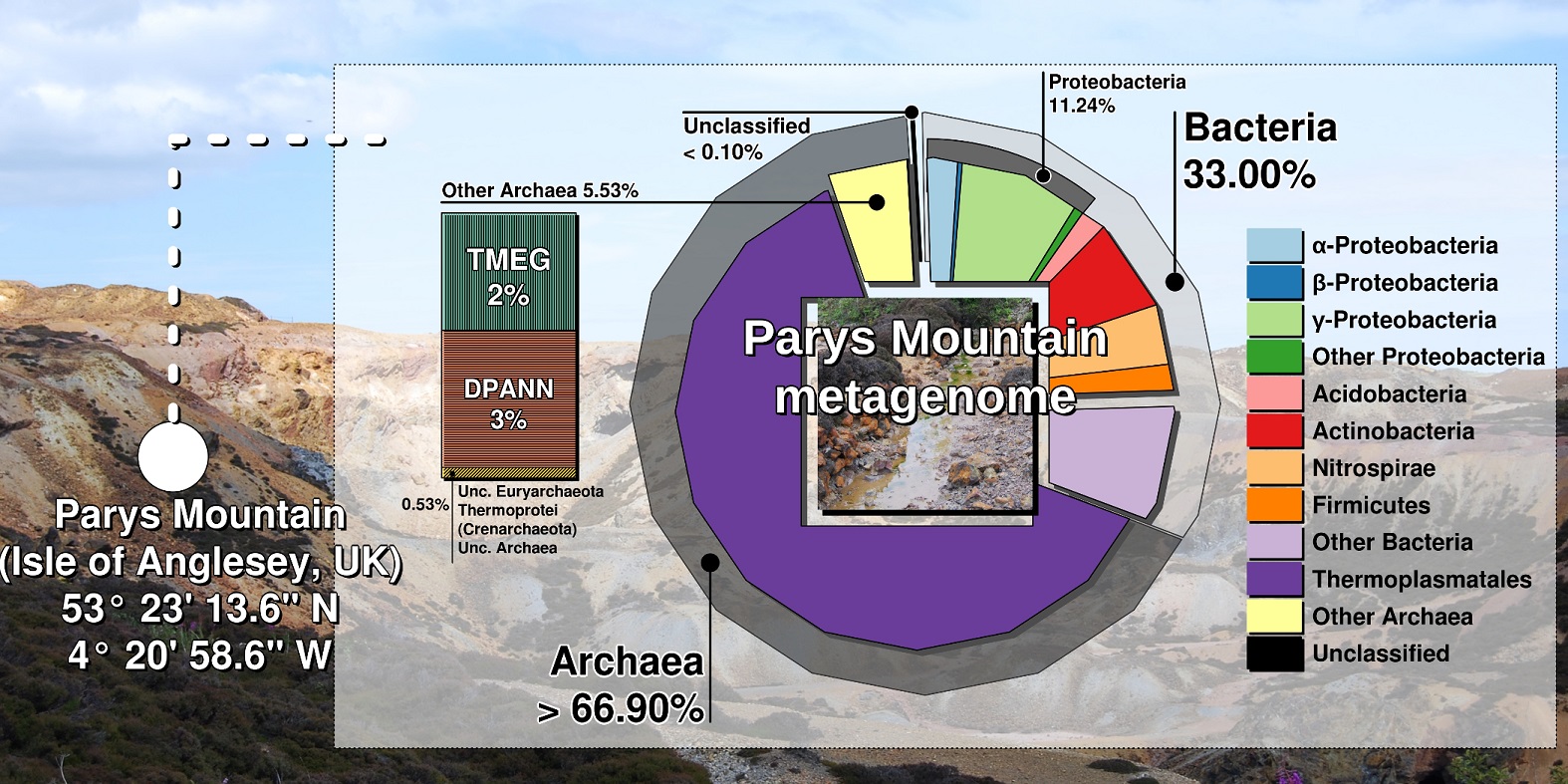
Among archaeal taxa, Cuniculiplasmataceae, “Candidatus Micrarchaeota” and “Terrestrial Miscellaneous Euryarchaeal Group” (TMEG) archaea were present in low numbers. TMEG archaea were earlier considered an “environmental clade” linked to Methanomassilicoccales-related archaea (4). Our global phylogenetic analysis of TMEG archaea showed strong clustering of Parys Mountain clonal variants with sequences detected in other acidic and moderately acidic places, pointing at the affiliation of this group with a new order within Thermoplasmata.
We also observed that this group of acidophilic organisms was often overlooked in acidic microbiomes. Highest numbers of archaea and in particular, Thermoplasmata were found in sediment samples, in contrast to the aqueous subsystem, where bacteria were in the majority. Furthermore, our analysis of microbial diversity in enrichment cultures pointed at the significance of sample pre-treatment, which could be used for isolation of cell wall-deficient archaea.
Overall, our results were counterintuitive: it is commonly accepted that acidophilic bacteria dominate low- and moderate-temperature environments, exemplified by Parys Mountain, which is exposed to the cool maritime climate. Many questions related to acidophilic Thermoplasmata still need to be addressed, e.g. the functions of predominant and “rare” community members and the reason of a very limited fraction of Thermoplasmata being cultured. Therefore, we are looking forward to sampling Parys Mountain in the near future.
1.Golyshina, O.V. et al. The novel extremely acidophilic, cell-wall-deficient archaeon Cuniculiplasma divulgatum gen. nov., sp. nov. represents a new family, Cuniculiplasmataceae fam. nov., of the order Thermoplasmatales. Int J Syst Evol Microbiol. 66(1), 332-40 (2016a).
2. Golyshina, O.V. et al. Biology of archaea from a novel family Cuniculiplasmataceae (Thermoplasmata) ubiquitous in hyperacidic environments. Sci Rep. 6, 39034 (2016b).
3. Golyshina, O.V. et al. “ARMAN” archaea depend on association with euryarchaeal host in culture and in situ. Nat Commun. 8, 60 (2017).
4. Söllinger, A. et al. Phylogenetic and genomic analysis of Methanomassiliicoccales in wetlands and animal intestinal tracts reveals clade-specific habitat preferences. FEMS Microbiol Ecol. 92(1), fiv149 (2016).
Follow the Topic
-
Microbiome
This journal hopes to integrate researchers with common scientific objectives across a broad cross-section of sub-disciplines within microbial ecology. It covers studies of microbiomes colonizing humans, animals, plants or the environment, both built and natural or manipulated, as in agriculture.

Related Collections
With Collections, you can get published faster and increase your visibility.
Oncobiome
This collection of papers delves into the burgeoning field of oncobiome research, exploring the intricate relationship between cancer and the microbiome. The oncobiome encompasses the diverse microbial communities residing in and on the human body, which influence cancer development, progression, and treatment responses. By examining these interactions, our aim is to unravel the complex mechanisms through which the microbiome impacts oncogenesis and therapeutic outcomes.
This compilation highlights cutting-edge research, offering insights into potential diagnostic markers and novel therapeutic strategies, thereby advancing our understanding of cancer biology and paving the way for innovative, microbiome-targeted cancer treatments.
This is a cross-journal collection between:
Experimental Hematology and Oncology
Articles will undergo the standard peer-review process of the journal to which they are submitted and are subject to either the BMC editorial policies or those of BJC Reports. Articles will be added to the Collection as they are published. The Editors have no competing interests with the submissions which they handle through the peer review process. The peer review of any submissions for which the Editors have competing interests is handled by another Editorial Board Member who has no competing interests.
Publishing Model: Open Access
Deadline: Ongoing
Animal Gut Nutrition and Greenhouse Gas Mitigation
Animal Microbiome, Journal of Animal Science and Biotechnology and Microbiome call for submissions to the collection on Animal Gut Nutrition and Greenhouse Gas Mitigation.
Efforts to reduce greenhouse gas emissions from livestock systems increasingly hinge on innovations in animal gut nutrition. The dynamic relationship between the gut microbiome and nutrient utilization plays a pivotal role in shaping methane output, feed efficiency, and overall sustainability. Advances in microbial ecology—particularly in understanding the role of gut microbiome in nutrient metabolism—are opening new pathways for mitigating emissions while enhancing productivity. These developments support the implementation of climate-smart agricultural strategies to address climate change and its impacts.
Looking ahead, continued research in this field has the potential to yield innovative solutions such as targeted probiotic supplementation, which could further optimize gut function and enhance nutrient absorption. These advancements may lead to reduced greenhouse gas emissions while improving animal health and productivity. By deepening our understanding of the animal gut microbiome, we can contribute significantly to sustainable agricultural practices that benefit both the environment and food security.
We invite researchers to contribute to this special Collection on Animal Gut Nutrition and Greenhouse Gas Mitigation. Topics of interest include but are not limited to:
- Animal Gut Microbiome and Feed Efficiency
- Greenhouse Gas Mitigation Strategies
- Rumen Fermentation Dynamics
- Nutrient Utilization in Livestock
- Probiotic Supplementation Effects
- Sustainable Livestock Production Practices
- Climate-Smart Agriculture Innovations
This Collection supports and amplifies research related to SDG 13, Climate action.
All submissions in this collection undergo the relevant journal’s standard peer review process. Similarly, all manuscripts authored by a Guest Editor(s) will be handled by the Editor-in-Chief of the relevant journal. As an open access publication, participating journals levy an article processing fee (Animal Microbiome fees, Journal of Animal Science and Biotechnology fees, Microbiome fees). We recognize that many key stakeholders may not have access to such resources and are committed to supporting participation in this issue wherever resources are a barrier. For more information about what support may be available, please visit OA funding and support, or email OAfundingpolicy@springernature.com or the Editor-in-Chief of the journal where the article is being submitted.
Publishing Model: Open Access
Deadline: Sep 04, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in