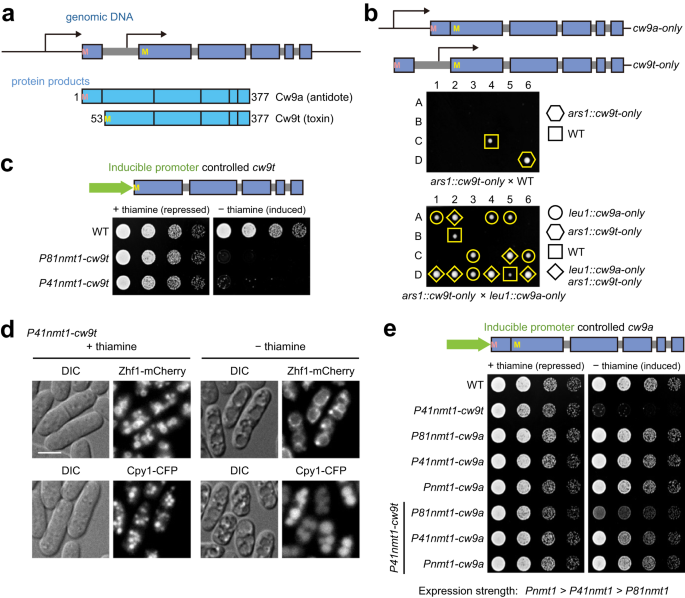
Mendel's law of segregation posits that in a diploid organism, each of the two alleles at a gene locus has an equal probability of being passed on to offspring. However, this law is disrupted by a type of selfish genetic element known as killer meiotic drivers (KMDs). KMDs disable non-carrier gametes, ensuring their transmission to over 50% of the viable offspring. Although KMDs have been found in animals, plants, and fungi, our understanding of their molecular mechanisms remains limited. Our recent Nature Communications paper unravels the workings of wtf-family KMDs, revealing how this family of single-gene KMDs can generate both a toxin and an antidote.
In 2016, when I started my PhD training at the National Institute of Biological Sciences in Beijing, China, I was introduced to the fantastic world of KMDs under the mentorship of Dr. Li-Lin Du and in collaboration with Dr. Wen Hu. Their knowledge and unwavering support were invaluable in my journey. It was a paradigm shift for me, as I had always been taught that genes are beneficial for organisms, but here were these selfish genes that benefit themselves at the expense of others.
The research on KMDs in the Du lab began with the study of within-species hybrid sterility in the fission yeast Schizosaccharomyces pombe. Dr. Hu noticed a low spore viability in the cross between the laboratory S. pombe strain and a wild S. pombe isolate CBS5557, indicating hybrid sterility. Pursuing the underlying cause of hybrid sterility, Dr. Hu identified two wtf (with Tf LTRs) family genes, cw9 and cw27, as single-gene KMDs (Hu et al., 2017). Concurrently, another wtf family KMD gene, wtf4 of the S. pombe var. kambucha strain, was discovered to produce both a toxin and an antidote from one gene (Nuckolls et al., 2017). These discoveries opened up an opportunity to apply the powerful fission yeast model to study the mechanisms of KMDs.
Intrigued by how the toxin and antidote of a wtf KMD operate, I embarked on a journey to unravel the killing and protecting mechanisms of cw9. We soon realized that conducting experiments in spores was challenging. To circumvent this difficulty, we decided to express the toxin (Cw9t) and antidote (Cw9a) separately in vegetative cells using inducible promoters. This strategy proved successful, with Cw9t exhibiting toxicity and Cw9a neutralizing this toxicity in vegetative cells.
The only difference between Cw9t and Cw9a is the extra 52 amino acids at the N terminus of Cw9a. Searching for physical interactors of this Cw9t-specific region led us to Rsp5/NEDD4 family E3 ligases, which bind and ubiquitinate Cw9a. The interaction is mediated by the PY motifs (Leu/Pro-Pro-X-Tyr) on the N terminus of Cw9a. Remarkably, by artificially altering the ubiquitination state, we could convert the toxin to an antidote and convert the antidote to a toxin. Ubiquitination of Cw9a promotes its endosomal transport and prevents Cw9a from becoming toxic. Protein-protein interactions between Cw9a and Cw9t result in the endosomal localization and detoxification of Cw9t.
Our journey was not without challenges. The toxicity of Cw9t and other toxic forms of Cw9 complicated strain construction, microscopic observation, and Western blotting analysis. Identifying the subcellular compartment where Cw9t localizes turned out to be a difficult endeavor. Genetic screening of ectopic mutations that can inhibit the toxicity of Cw9t failed to yield informative clues. These obstacles hindered our investigation into the killing mechanism of Cw9t.
We also encountered challenges with Cw9a. Initially, we used the C-terminally tagged Cw9a for various experiments. However, we found that the C-terminal tag can inhibit the toxicity of Cw9a with PY motif mutations, so we had to switch to use N-terminally tagged Cw9a. Another issue was degradation under non-denaturing conditions. We had to employ vacuolar protease mutant background to prevent Cw9a degradation. For in vitro ubiquitination assay, initially the unreacted substrate band was only visible by gel staining, not Western blotting (Picture 1). After much trial and error, we solved the problem by using glutaraldehyde fixation to prevent protein loss from blotted membrane, enabling the Western blot-based detection of the unreacted protein.
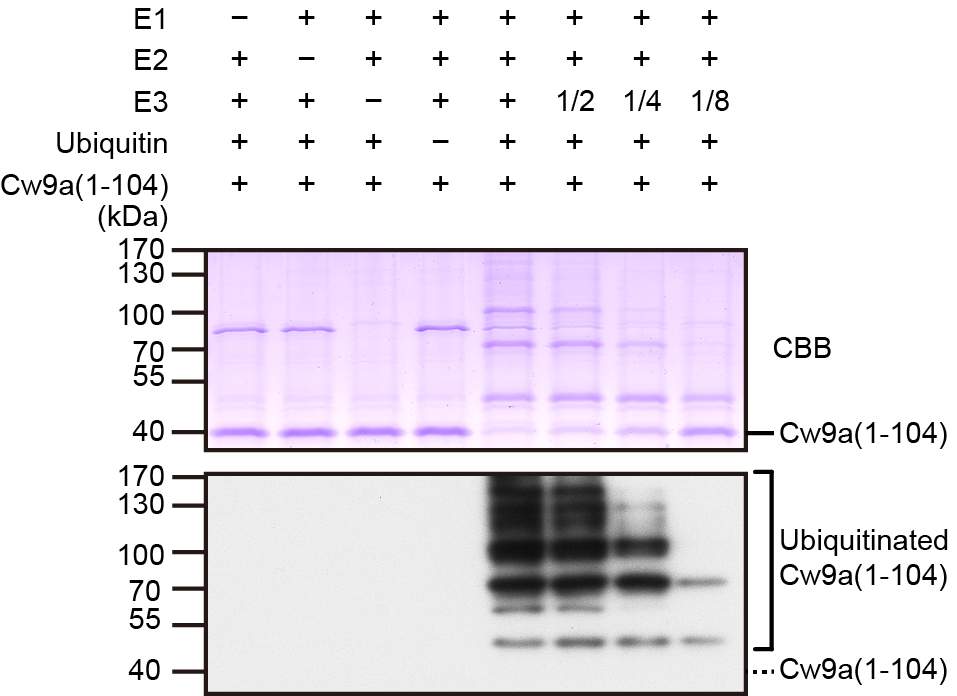
Picture 1. Pub1-catalyzed in vitro ubiquitination of Cw9a(1-104). The band of unreacted 8×myc*-Cw9a(1-104) is indicated in the results of Coomassie brilliant blue (CBB) stained gel and Western blot with myc antibody. The band of Cw9a(1-104) was not visible by Western blot.
Despite challenges, my six-year journey unraveling the toxin-antidote duality of cw9 has been rewarding. I am grateful for the enriching experiences and the invaluable assistance I have received throughout this journey. It is my earnest hope that this work will contribute to a broader understanding of KMDs and pave the way for future discoveries in this fascinating field.
References
Hu, W., Jiang, Z. D., Suo, F., Zheng, J. X., He, W. Z., and Du, L. L. (2017). A large gene family in fission yeast encodes spore killers that subvert Mendel’s law. eLife, 6, e26057.
Nuckolls, N. L., Bravo Núñez, M. A., Eickbush, M. T., Young, J. M., Lange, J. J., Yu, J. S., Smith, G. R., Jaspersen, S. L., Malik, H. S., and Zanders, S. E. (2017). wtf genes are prolific dual poison-antidote meiotic drivers. eLife, 6, e26033.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in