You’re not gonna need a bigger jaw
Published in Ecology & Evolution


Modern sharks are a diverse and globally distributed group of marine vertebrates whose fossil record can be traced back to the Early Jurassic (ca. 184 Mya), and since that time they have developed a wide range of lifestyles and expanded into various ecological niches. However, the evolutionary trajectories leading to today’s shark diversity are still poorly understood.
I have been studying sharks for nearly six years since starting my doctoral studies at the University of Vienna in Austria. Sharks represent a great opportunity to understand how predators have evolved, and how this is reflected in their morphology. The fossil record of sharks is mainly dominated by their teeth, as to be expected since teeth are more likely to be fossilized, as opposed to their cartilaginous endoskeletons. However, some shark body plans have remained consistent through their evolutionary history (Lopez-Romero et al, 2020). Thanks to a great collaboration with tomographic data of sharks (Figure 1) (Kamminga et al, 2017, Dearden et al 2021), studying the morphological evolution of sharks is now catching up with other groups that have been more thoroughly studied.

Besides teeth, the feeding apparatus’ are a good proxy to infer several morpho-functional traits, not only for sharks, but also for many jawed vertebrate (Deadkin et al, 2022). Its relevance shows that there is a link between the prey capture and bite force which has been extensively studied in sharks’ biomechanics and its relationship with the cartilage that forms the jaws. Interestingly, the jaws of sharks are quite unique among extant vertebrates. On the one hand, they are developed from a single structure, called the Meckel’s cartilage and the palatoquadrate. These elements are also found in other vertebrates, but only during their embryonic development as a transient structure which eventually is replaced by dermal and endochondral bone. This facilitates somehow the recognition of possible homologies and thus the comparison to other vertebrate groups. However, since the sharks’ mandibular arch is composed of only two elements, the precise recognition of discrete locationscan be somehow tricky. This is important since we studied the jaw evolution thorough geometric morphometrics, which is a way to statistically analyze shapes (leaving outside differences in size, rotation and orientation). To perform these analyses, it is necessary to recognize homologous locations (landmarks) (Figure 2). And while we were able to find some equally comparable landmarks, other aspects of the jaw were also relevant. As seen in the figure, the posterior part of the jaw, where it articulates to the palatoquadrate, provides a distinct insertion point for muscles, and as such bears information on differences derived from feeding. The feeding mode of sharks have been subject to numerous studies and different evolutionary pathways have been proposed (Moss, 1977; Wilga et al, 2007). However, our understanding the shark interrelationships has changed considerably over the years. For instance, morphological phylogenies recovered rays and skates as highly derived sharks. This scheme was widely used in previous works describing the evolution of jaw suspension among sharks, which was thought to yield a phylogenetic signal. Later phylogenies using molecular data challenged this view and confirmed that skates and rays actually form the sister group of sharks. For our project, we used the phylogenetic data of Stein et al (2018), which covers all extant shark orders comprising 511 species.

The project involved a large team of colleagues since its initial conceptualization under the leadership of Dr Martin D. Brazeau. The X-ray computed tomography database used contains 95 shark species (later some were removed due to difficulties in placing landmarks) that cover all major shark orders, and served as a starting point for studying the morphological evolution of sharks. One of the first results of the initial 3D geometric morphometrics analysis showed a fairly consistent separation of the major orders and, to some extent, limited variation among some of the most speciose orders (requiem sharks), while other groups showed a divergent pattern as in the carpet sharks. Most interesting was the finding that one of the Squaliformes, which occurs mainly in deep waters, had greater variation in its jaws than the rest of the orders. This prompted us to further investigate what factors might influence this pattern and how well this is reflected in the morphological differences of the other orders as well as in the morphological evolutionary rates.
The next step was to gather all possible information that could explain some of the differences we observed. With this in mind, some initial clues were related to prey content. Due to the importance of sharks as predators in their habitat and their importance for fisheries, several surveys on the stomach contents of many species are available for most of the species included in the study. Of course, these data are subject to many variations depending on the age of the individuals surveyed and the location where the individuals were caught, in addition the information is updated for many species. For this reason, we have also included trophic level information, which may include larger groups regardless of their specific stomach contents. These factors have been shown to covary with other traits, such as size in sharks, which has been used to study evolution towards large, high-level predators. Another variable that has been shown to influence speciation rates in sharks is habitat. Several species occur near reefs, and this pattern is consistent in bony fishes, as well as other benthic species, even in fossil records (Sorenson et al, 2014). Another important recent discovery in bony fishes is that deep-sea species tend to show greater morphological disparity. Among sharks in particular, the order Squaliformes is mainly distributed in deep waters. This was described around the Cretaceous period, when they colonised the deep sea. This event was also followed by high speciation rates in some of the families, along with other morphological changes such as the presence of bioluminescence (Duchatelet et al, 2021).
One of the most important results of our analysis was the finding of a decoupling between evolutionary rates and morphological disparity in some of the orders studied. Most of the species distributed on the reef show a lower overall disparity and morphological evolution rate than the deep-sea group. This confirms previous studies that the deep sea is an important habitat where very diverse forms occur (see Martinez et al, 2021). And not only has the colonisation of the deep sea by sharks been accompanied by the appearance of bioluminescence, but also by other morphological changes that can be observed in their jaws. Other aspects of shape also reflect biomechanical properties related to primary feeding strategies. Finally, when we examined disparity over time (Figure 4), we found that the morphological evolution of the shark jaw appears to follow an early burst model, which may explain the reduced disparity seen in some groups today. Furthermore, as in palaeodiversity studies, we found evidence for an increase in disparity around the mid-Cretaceous. Our study serves as a starting point for research into other aspects of shark morphological evolution. Many improvements can be made when fossils are integrated into studies like ours. Although it is difficult to obtain a well-preserved shark fossil that retains its shape, some specimens have the potential to be included and provide a refined picture of the evolutionary rates we observed.
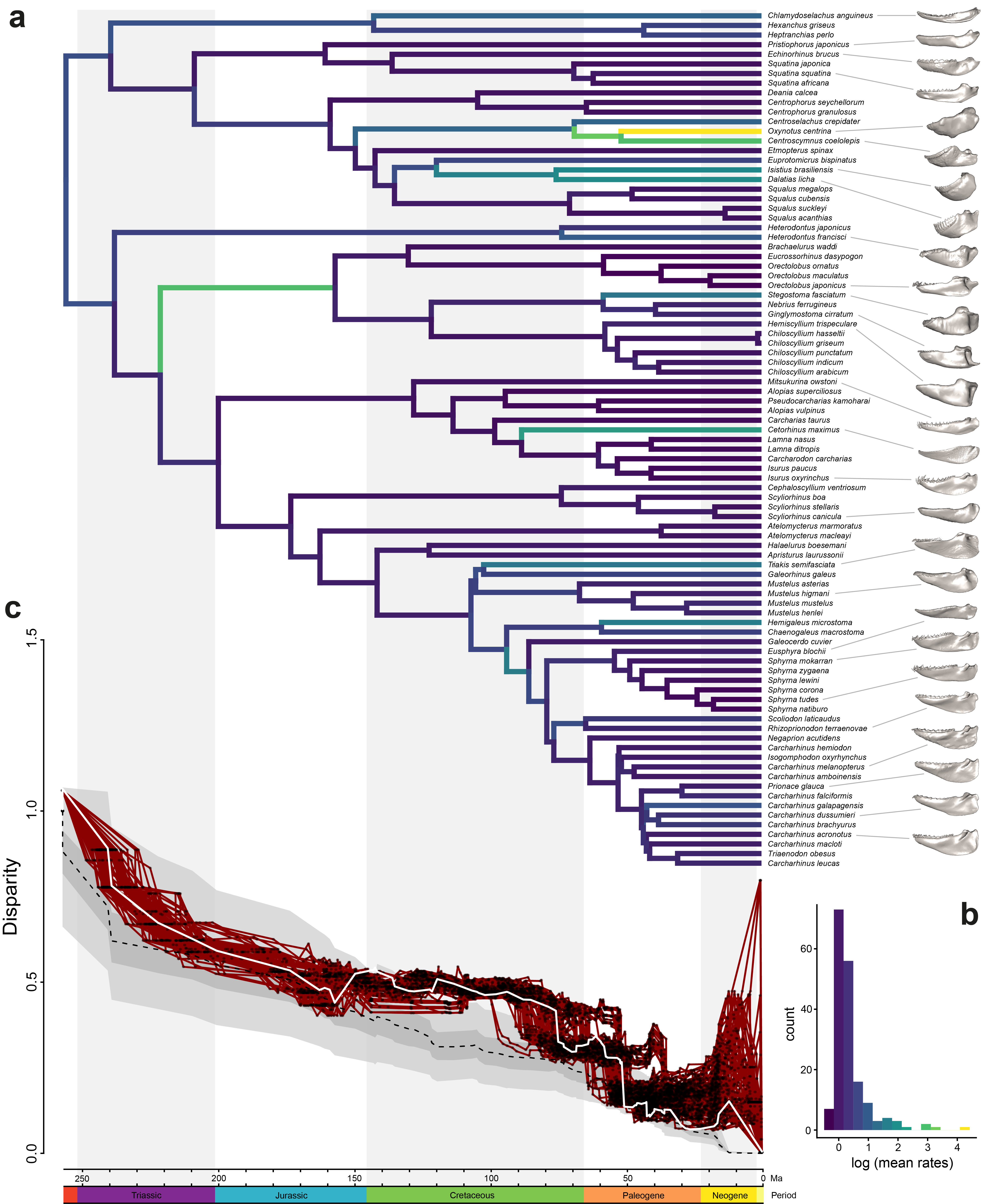
Warmer colors in the phylogeny shows higher rates, and lower plot shows the disparity among the groups through time.
Note: Poster image is a close up of Dalatias licha (kitefin shark). Image credit: Jürgen Kriwet.
References:
Deakin, W. J., Anderson, P. S., den Boer, W., Smith, T. J., Hill, J. J., Rücklin, M., ... & Rayfield, E. J. (2022). Increasing morphological disparity and decreasing optimality for jaw speed and strength during the radiation of jawed vertebrates. Science Advances, 8(11), eabl3644.
López-Romero, F. A., Stumpf, S., Pfaff, C., Marramà, G., Johanson, Z., & Kriwet, J. (2020). Evolutionary trends of the conserved neurocranium shape in angel sharks (Squatiniformes, Elasmobranchii). Scientific Reports, 10(1), 12582.
Kamminga, P., De Bruin, P. W., Geleijns, J., & Brazeau, M. D. (2017). X-ray computed tomography library of shark anatomy and lower jaw surface models. Scientific Data, 4(1), 1-6.
Dearden, R. P., Mansuit, R., Cuckovic, A., Herrel, A., Didier, D., Tafforeau, P., & Pradel, A. (2021). The morphology and evolution of chondrichthyan cranial muscles: A digital dissection of the elephantfish Callorhinchus milii and the catshark Scyliorhinus canicula. Journal of Anatomy, 238(5), 1082-1105.
Moss, S. (1977). Feeding mechanisms in sharks. American Zoologist, 17(2), 355-364.
Wilga, C. D., Motta, P. J., & Sanford, C. P. (2007). Evolution and ecology of feeding in elasmobranchs. Integrative and Comparative Biology, 47(1), 55-69.
Stein, R. W., Mull, C. G., Kuhn, T. S., Aschliman, N. C., Davidson, L. N., Joy, J. B., ... & Mooers, A. O. (2018). Global priorities for conserving the evolutionary history of sharks, rays and chimaeras. Nature ecology & evolution, 2(2), 288-298.
Sorenson, L., Santini, F., & Alfaro, M. E. (2014). The effect of habitat on modern shark diversification. Journal of Evolutionary Biology, 27(8), 1536-1548.
Duchatelet, L., Claes, J. M., Delroisse, J., Flammang, P., & Mallefet, J. (2021). Glow on sharks: state of the art on bioluminescence research. Oceans, 2(4), 822-842.
Martinez, C. M., Friedman, S. T., Corn, K. A., Larouche, O., Price, S. A., & Wainwright, P. C. (2021). The deep sea is a hot spot of fish body shape evolution. Ecology letters, 24(9), 1788-1799.
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026
DNA repair and human disease
Publishing Model: Hybrid
Deadline: Oct 31, 2026



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in