15 non cross-reacting protein glues for one-pot seamless assembly of multiple peptides
Published in Bioengineering & Biotechnology
By Filipe Pinto and Ella Lucille Thornton
Nature has evolved numerous proteins for the manipulation of DNA, which we use in the lab on a daily basis. We cut it, ligate it, modify it; but what about proteins? Of course, there are proteases that can cut proteins, but tools for the post-translational ligation of proteins are somewhat more unusual. Here, we show the remarkable capabilities of split inteins as protein ligation tools (Fig. 1).
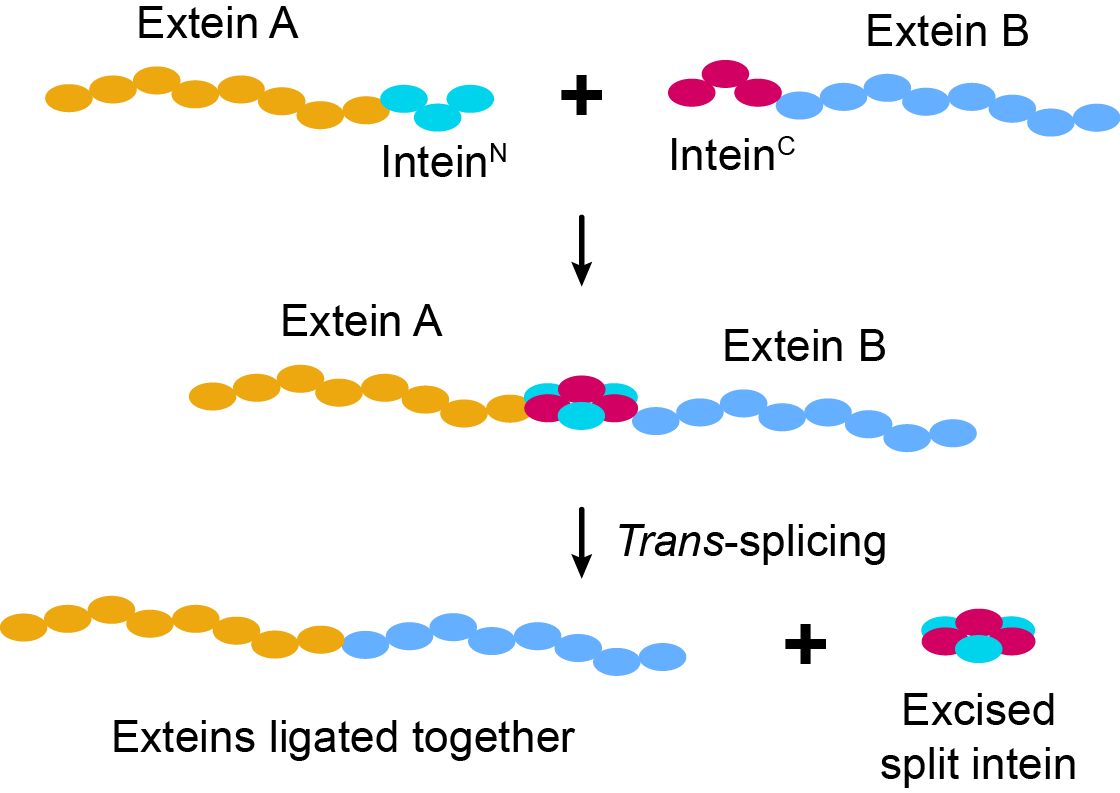
Figure 1 | Protein ligation using split inteins.
Inteins are protein segments capable of protein ligation and have been referred to as “Nature's gift to protein chemists”1. The process of joining adjacent residues via a peptide bond is known as protein splicing (in trans if the intein is split) and the intein itself is not present in the final sequence, thus achieving scarless peptide ligation.
The self-catalytic nature of the splicing reaction, which relies solely on protein folding and the fact that the inteins are not part of the final product, are characteristics of exceptional value; thus, making them remarkable tools for both protein engineering and synthetic biology applications.
As nothing is flawless, inteins have particular preferences for the amino acid residues on either side of the ligation site. Deviation from these 6-residue sequences (3 aa. on each extein) may affect splicing efficiency, thus making life difficult for those wishing to use inteins with proteins that cannot tolerate amino acid residue additions or substitutions.
The more you have, the more you can choose from!
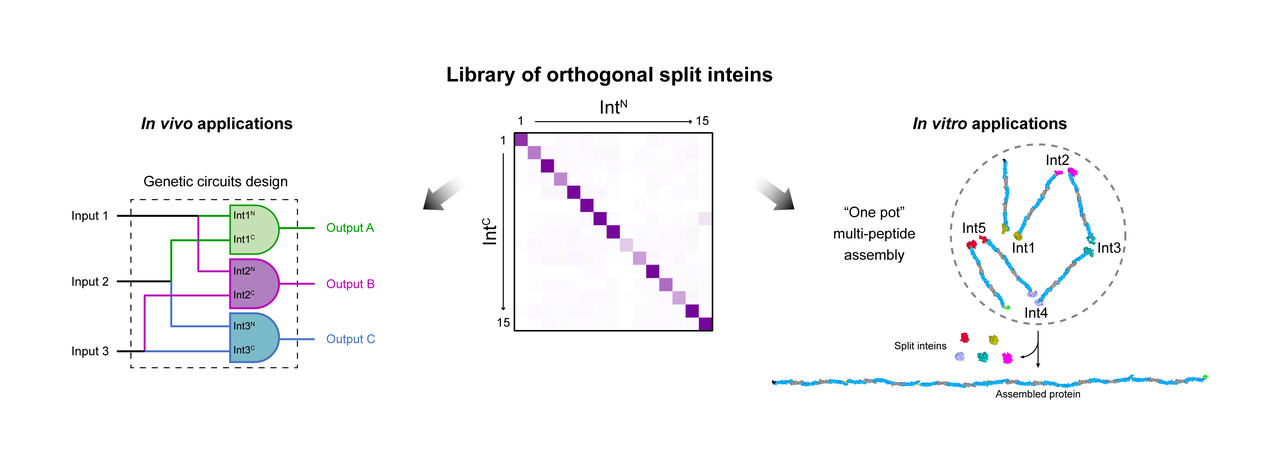
Bearing this in mind, we aimed to expand the set of orthogonal inteins currently available, to increase the chances of finding inteins compatible with the target proteins and to be able to use them simultaneously. To this end, we searched the literature and InBase2 (the intein database) for inteins with different native insertion sites (exteins) and selected an initial set of 34 phylogenetically distant inteins (Fig. 2), with the assumption that homology negatively correlates with orthogonality.
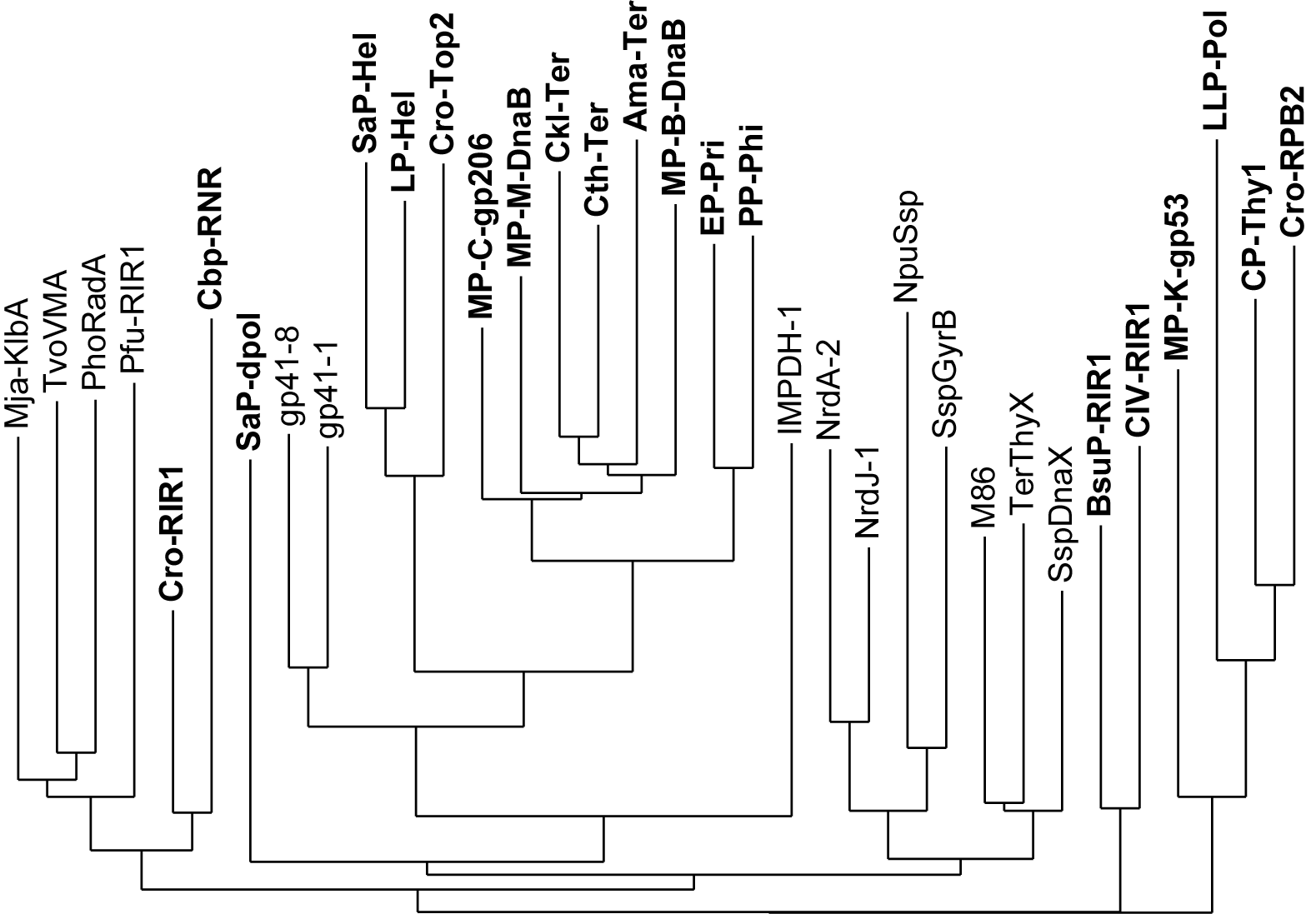
Figure 2 | Phylogram based on the structural alignment of the 34 inteins selected for characterization (inteins retrieved from InBase are in bold).
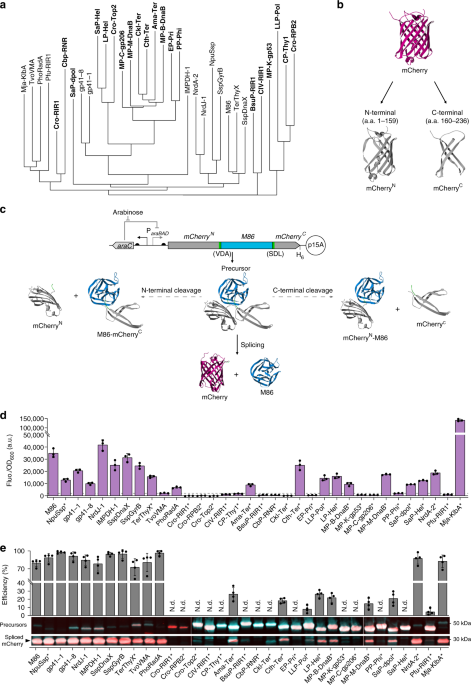
Next, came the question of how to efficiently characterize such a large library of inteins under similar conditions and close to their native context. To address this, we split the fluorescent protein mCherry and observed how efficiently and quickly our library of split inteins was able to glue back together the two halves and reconstitute fluorescence (Fig. 3a). Not only did this approach allow us to analyse how each intein performs, it also enabled a high-throughput technique to screen for cross-reactivity between the inteins (Fig. 3b). We were able to identify 15 highly orthogonal split inteins, five of which were previously uncharacterised, and defined reaction conditions in which 10 of them can be used concurrently in vitro.
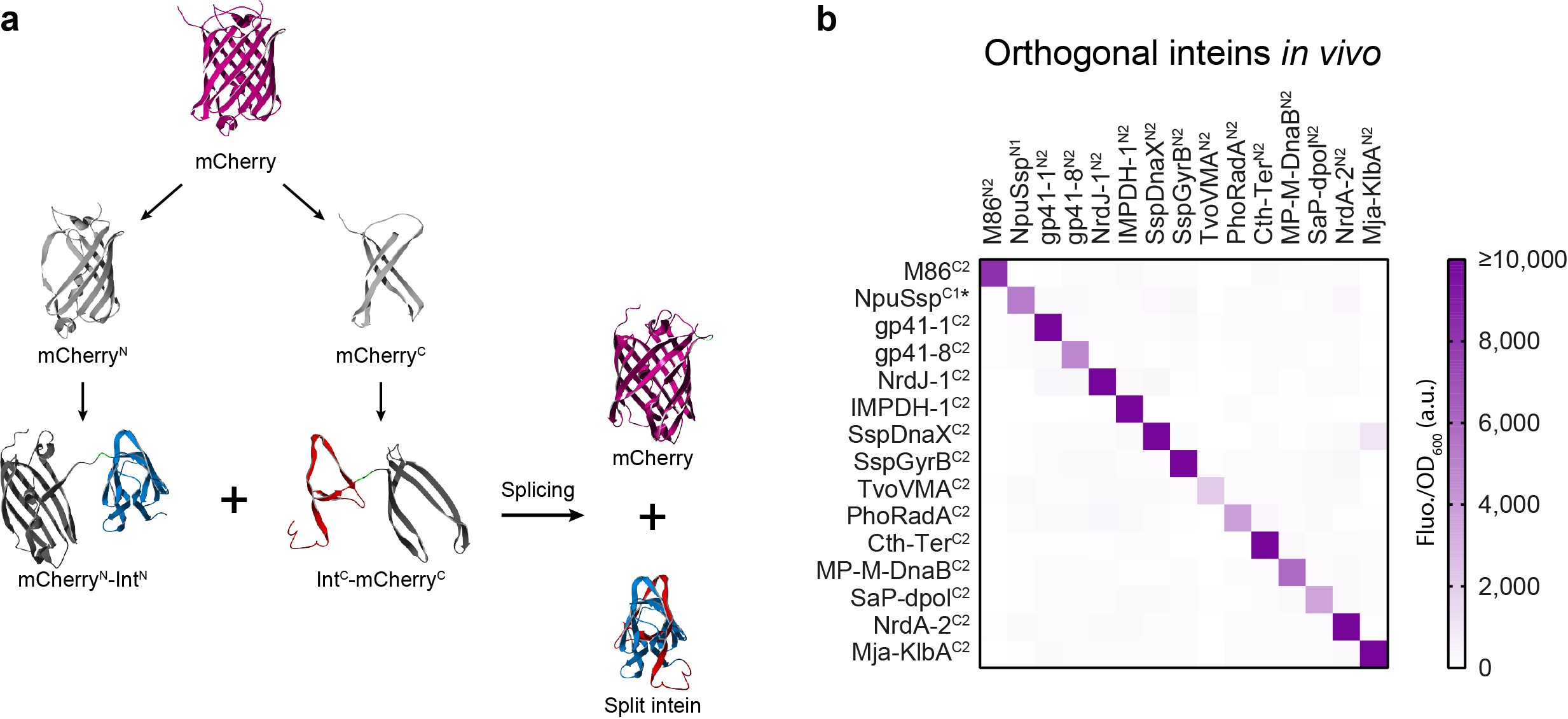
Figure 3 | A novel split mCherry platform (a) for rapidly assessing split inteins trans-splicing and orthogonality (b).
“Once you have the tools to build your dreams, they are a step closer to become a reality!” FP
With this expanded library in hand, we further demonstrated its versatility in enabling new synthetic biology (in vivo complex logic circuit design) and protein engineering (in vitro modular protein assembly) applications.
(i) We have built and connected three orthogonal AND gates based on intein-split extracytoplasmic function (ECF) sigma factors to develop a 3-input 3-output integrated logic circuit (Fig. 4a). It produces fluorescent outputs when Escherichia coli cells are exposed to two or more inputs and discriminates between them by reporting a corresponding fluorescent protein.
(ii) We have performed the in vitro assembly of a large (~226 kDa) highly repetitive protein from smaller precursors individually expressed in different bacterial cells. We directly used clear E. coli lysates (no precursors’ purification required) to assemble the protein in ‘one pot’ using 5 orthogonal inteins (Fig. 4b) or by on column assembly, using only two inteins to glue the protein fragments one after the other, reusing cell lysates.
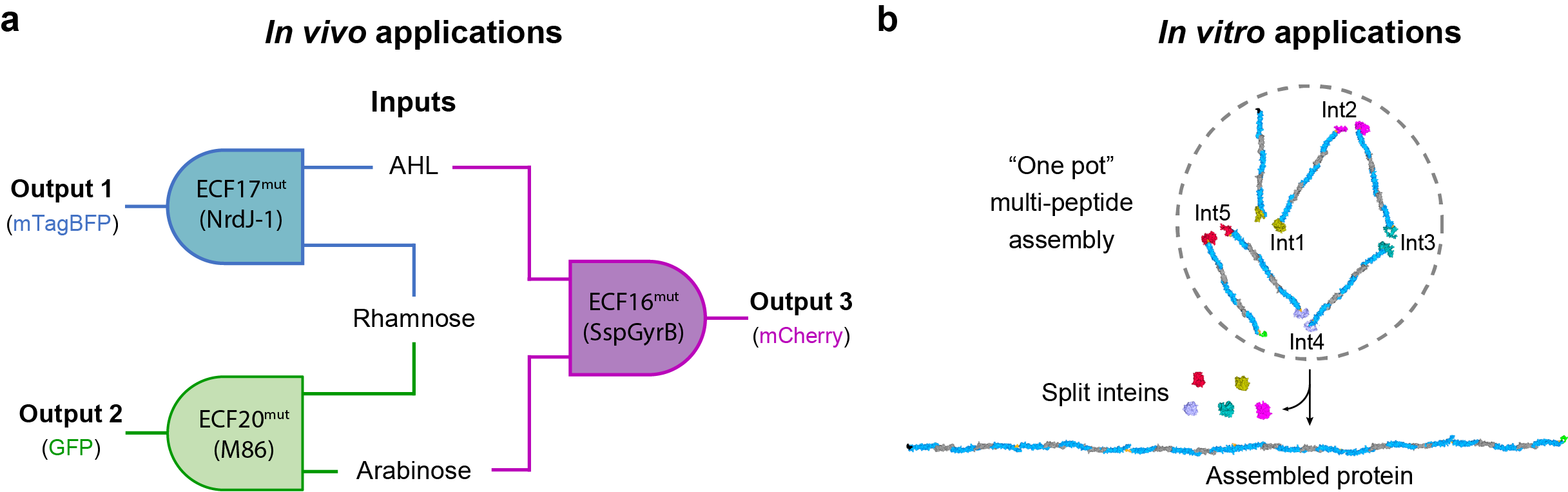
Figure 4 | Split inteins enabled complex genetic circuits (a) and in vitro large protein assemblies (b).
This work provides an expanded library of orthogonal split inteins and shows its vast potential in both in vivo applications, including scaling up genetic logic circuit design in living cells, and in vitro applications, including providing a rapid and simple way to seamlessly assemble large repetitive proteins that are of biotechnological interests but often difficult to clone and produce using heterologous expression systems.
We believe that our work will be of interest to the broad scientific community, in particular to synthetic and chemical biology researchers who seek new enabling tools to scale up gene circuit design and new efficient methods for protein assembly and biomaterial engineering.
The paper:
Pinto F, Thornton EL & Wang B. An expanded library of orthogonal split inteins enables modular multi-peptide assemblies. Nature Communications, 11, 1529 (2020). DOI: 10.1038/s41467-020-15272-2
References
1. Shah NH & Muir TW. Inteins: nature’s gift to protein chemists. Chem. Sci. 5, 446-61 (2014).
2. Perler FB. InBase: the Intein Database. Nucleic Acids Res. 30, 383-84 (2002).
Filipe Pinto (born 1981) has a first degree (2003) and an MSc (2009) in Biochemistry, and a PhD in Biology in the field of Synthetic Biology (2013, Approved with Distinction), all from the University of Porto (UP) in Portugal. He did an Erasmus internship at BRC (Uni. Szeged, HU) in his final college year and since then he was involved in research activities in the fields of molecular microbiology and protein science. He studied phage-triggered bacterial autolysis (2004, Uni. Lisbon, PT), integrated a group of molecular structure of proteins (2005, IBMC/UP, PT) and from 2006 to 2015 he focused his research in the molecular biology of cyanobacteria (IBMC/UP, PT). During this period, he integrated the research team led by Paula Tamagnini and participated in two Synthetic Biology European projects: the pioneer FP6 EU-project “BioModularH2” as a PhD student, and the FP7 EU-project “CyanoFactory” as a Postdoc. He was also involved in the preparation of proposals of funded R&D projects and was part of the research team performing the work; he contributed to the creation of the ideas as well as in writing the proposals. From 2016 to 2021, he was a Postdoctoral Research Associate in the field of Synthetic Biology at the University Edinburgh (UK), accumulating lab manager functions. During this period, his research interests included the design and engineering of new scalable biological circuits by developing and combining libraries of orthogonal split inteins (“protein ligases”), optimized extracytoplasmic function (ECF) sigma factors and orthogonal transcription repressors.
Currently, F Pinto is an Assistant Researcher (CEEC- Individual Call – 3rd Edition) at the Bioengineering & Synthetic Microbiology group (i3S/UP, PT), developing his research in the field of cyanobacterial Synthetic Biology.
With 15+ years of research experience, F Pinto co-authored his first paper while in his degree and, in total, he published 18 papers (4 of which as lead author and 1 as co-first author) in well-established international peer-reviewed scientific journals (with an accumulated IF of over 120), with 560+ citations and h-index 11 (Scopus Author ID: 35614368600). Six of the papers were published in the last 5 years and he was lead author in one publication that resulted in granted European patents. The work he developed in this period also resulted in 3 research articles published in Nature Communications, a highly regarded peer-reviewed journal. He was co-guest editor in a Synthetic Biology special issue in the MDPI journal Life (ISSN 2075-1729). He participated in several national and international meetings where he presented 4 oral communications and 29+ conference posters, including two prestigious Gordon Research Conferences on Synthetic Biology.
Regarding F Pinto’s pedagogical role, he co-supervised an undergraduate student within the FP7 EU-project and two MSc students at UoE, and he was involved in the supervision of several other students, including high-school, undergraduate (including 2 Erasmus), MSc and PhD students. He was also one of the coordinators of the RT-qPCR course, part of the advanced training provided by the Doctoral Program in Molecular and Cell Biology (IBMC, ICBAS, FCUP; UP) in the academic years of 2013-14 and 2014-15. From 2009 to 2015, he gave invited talks in the Environmental Biotechnology syllabus from the Master in Ecology, Environment and Territory (FCUP) and the integrated Master in Bioengineering (FEUP). From 2010 to 2015, F Pinto was part of “Embaixadores da Ciência”, a group of IBMC/INEB researchers engaged in science dissemination in schools.
He was awarded a 1st poster prize and 4 conference travel awards, and the prize “Prémio Fundação Eng. António de Almeida” that distinguishes the best FCUP PhD thesis in the areas of Life Sciences, Agricultural Sciences and Landscape Architecture. He is inventor in recently granted patents describing the neutral sites technology in the cyanobacterium Synechocystis (European Patents EP3106521 & EP3461897).
Follow the Topic
-
Nature Communications

An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.
Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Biosensing
Publishing Model: Hybrid
Deadline: Jun 30, 2026



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in