Can a herpesvirus help us fight rabies in vampire bats?
Published in Microbiology

The blood feeding behaviour of common vampire bats is the perfect mechanism for the saliva-based transmission of Rabies virus. Despite the increasing health and economic impact across Central and South America, few strategies aside from mass culls exist to combat vampire bat rabies.
My lab group and numerous collaborators have been investigating ways in which the current management of rabies virus in vampire bats can be improved, including the vaccination of bats. This is an enormous challenge for a couple of main reasons:
- Wildlife vaccines are usually distributed in edible baits; however, the diet of vampire bats means that they can’t be vaccinated in this way.
- An alternate method of vaccination is capture, injection and release. It would be impossible and horribly expensive to achieve a meaningful proportion of bats, given the many hundreds of thousands across such a large and inaccessible area.
An ideal method of vaccination would be one that spreads between bats on its own, without further input: a transmissible vaccine. One method of creating such a vaccine would be to engineer a harmless virus of the host species of interest (vampire bats) to express the rabies virus glycoprotein. This virus would then vaccinate those it infected against rabies, and spread unaided through the population.
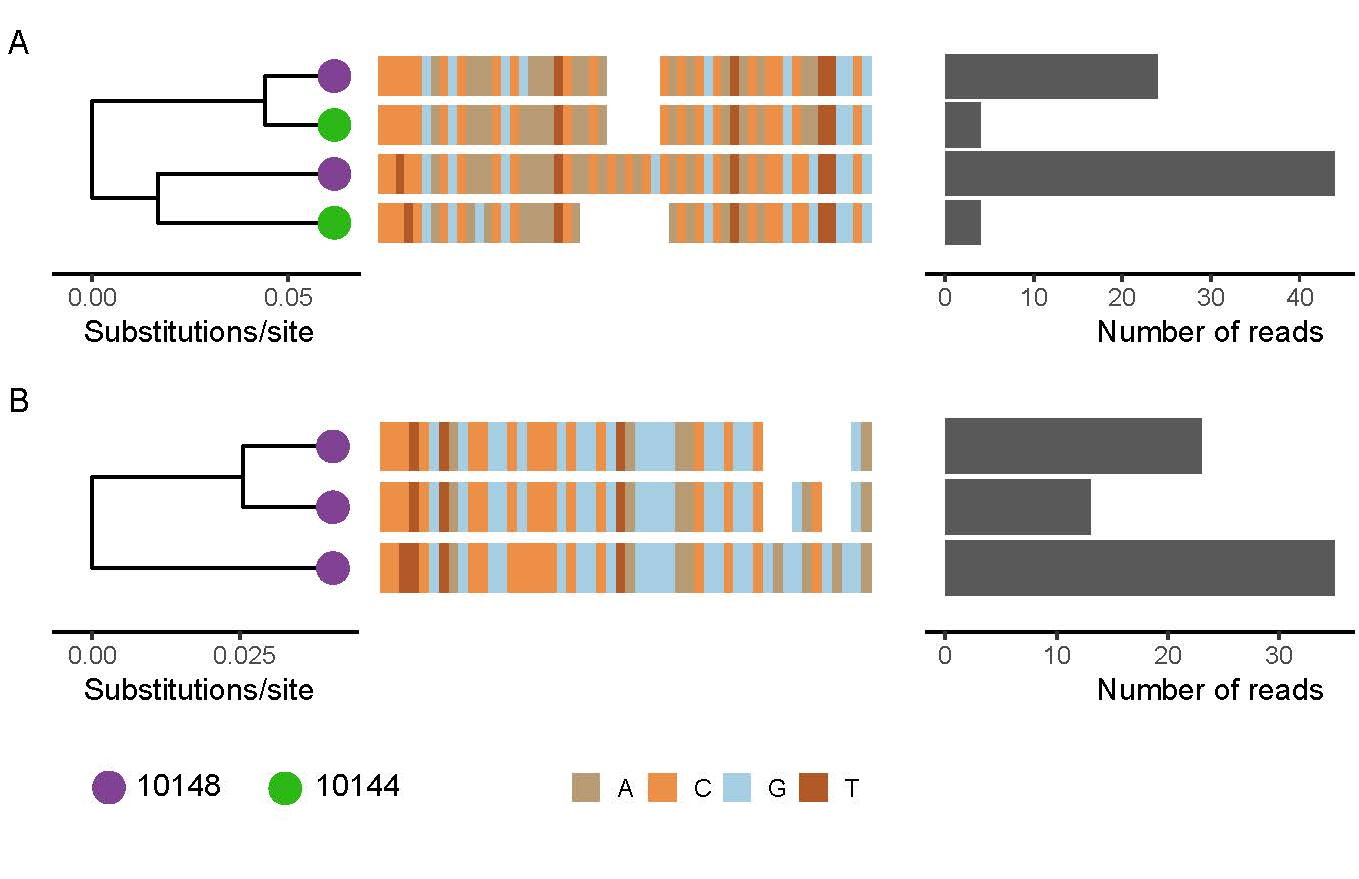
We set out to investigate whether a betaherpesvirus discovered in a vampire bat saliva sample through metagenomic sequencing would make a good candidate for further study as a transmissible vaccine vector for rabies virus. At our disposal were several years’ worth of field samples from vampire bats. Using a betaherpesvirus PCR on over 100 saliva samples, we found a promisingly high prevalence of 96%, which suggests that as a vaccine vector, it would be able to spread to a high level of coverage. We also tested the few saliva samples from other Peruvian bat species that we had available, to compare how closely related their herpesviruses were. The viruses formed largely species-specific clades, apart from one Artibeus lituratus bat sample which carried a virus identical to that of the vampire bats. Unfortunately, our sample size for this species is so small (N=2) that we could not confirm if this was a one-off cross transmission event, or if the virus can circulate between the closely related species. Nevertheless, it appears that there would be no transmission outside of this bat family, especially not to livestock or humans.
We also picked two herpes-positive saliva samples to undergo metagenomic sequencing, in order to get a complete genome for the herpesvirus. During the process of whole-genome reconstruction from sequence reads, it became clear that in a single sample, more than one strain of virus was present. This was both good and bad news.
The good: a multiple strain infection strongly implies that super-infection by a different strain of virus can take place after initial infection, a process absolutely crucial for a vaccine vector being introduced into a heavily wild-type infected population. The bad: multiple strains meant that it was impossible to reconstruct a single whole genome sequence, as the short (150bp) reads could not be separated into separate strains. As such, the genome we constructed was a chimera of the different strains present (the ugly!).
In summary, using saliva samples taken from wild vampire bats, we identified a promising candidate betaherpesvirus for continued investigation as a transmissible vaccine vector against rabies virus. We hope to continue this work and further characterise the betaherpesvirus.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in