Glucocorticoid resistance in B cell acute lymphoblastic leukemia
Published in Cancer
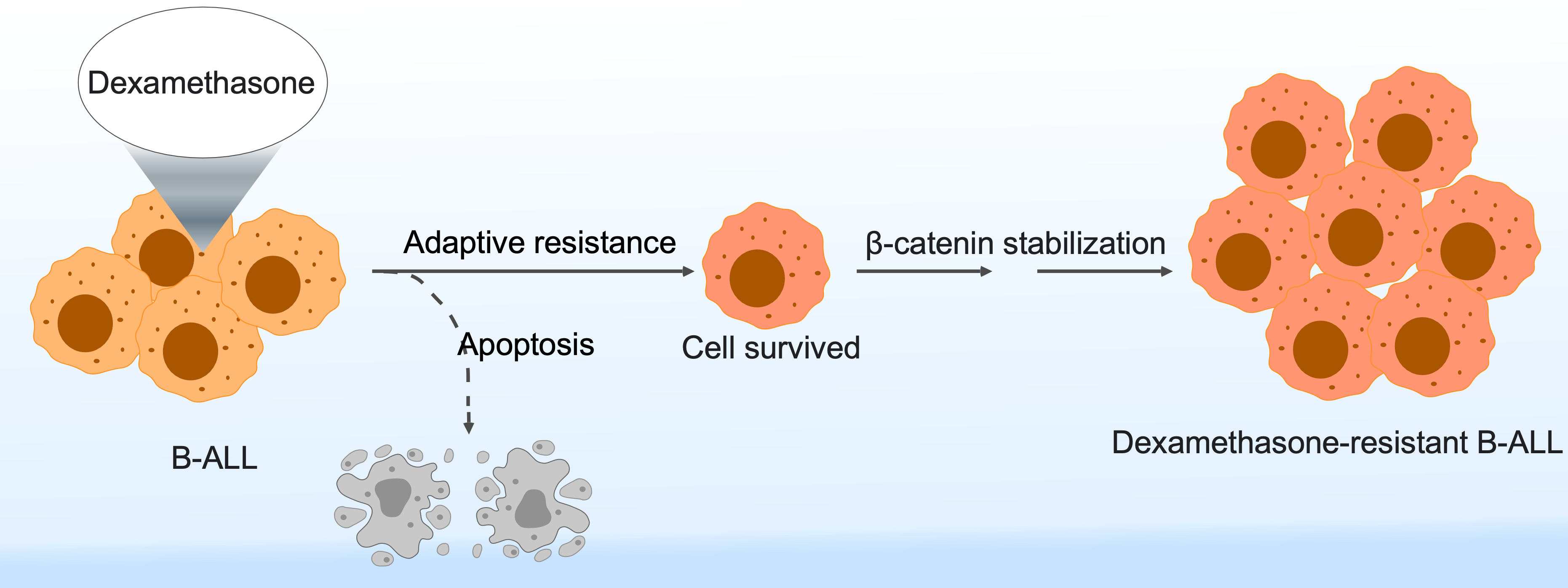

Despite incredible progress in anticancer therapy development, resistance to therapy is the major factor limiting the cure of cancer patients. It results in up to 90% of cancer-related deaths in various types of cancer.1 Therefore, identification of the factors that drive the development of therapy resistance is a pressing issue in the field.
Several mechanisms explaining the development of therapy resistance have been proposed.2 The most studied mechanism is the so-called Darwinian evolution, which assumes a selection of clones harboring rare pre-existing genetic alterations or the acquisition of de novo mutations, which then survive therapy-induced death. Another mechanism is Lamarckian selection, which assumes that therapy induces epigenetic modifications, leading to alterations in the expression of tumor suppressors or oncogenes, with an ultimate activation of cell survival pathways.2 Dynamic fluctuation has recently been proposed as yet another mechanism. It assumes that dynamic changes in gene expression in some cells allow these cells to escape therapy-induced death.3 However, in many cases, the specific molecular events governing the development of therapy resistance remain largely unknown.
In 2017, we set a goal of identifying glucocorticoid-mediated transcriptomic changes in acute lymphoblastic leukemia (ALL). Glucocorticoids such as dexamethasone and prednisolone are widely used in the treatment of ALL.4 We treated several ALL cell lines with dexamethasone and prednisolone and analyzed them for transcriptional changes. We observed that while dexamethasone and prednisolone induced apoptosis signaling in glucocorticoid sensitive ALL cells, several oncogenic pathways including the β-catenin stabilization pathway were enriched in both dexamethasone and prednisolone-treated cells. This finding was interesting and we hypothesized that glucocorticoid-treatment might influence the core cellular signaling network. Protein kinases play important roles in core cellular signaling.5 Therefore, we planned to check whether glucocorticoid treatment alters the activity of protein tyrosine and serine/threonine kinases. We used Pamgene peptide substrate-based kinase profiling to identify protein tyrosine and serine/threonine kinase activity after dexamethasone treatment. We observed that while tyrosine kinase activity was abolished in dexamethasone-treated cells, the activity of several serine/threonine kinases including aurora kinase, S6K, etc. was enhanced. We further demonstrated that dexamethasone treatment enhanced p38, S6K, and GSK3β phosphorylation as well as β-catenin stabilization over time.
Next, we wanted to check whether ALL patient resistance to dexamethasone displays similar pathway enrichment. We developed a deep learning model to detect dexamethasone sensitivity in ALL and used the model to predict dexamethasone sensitivity in ALL. While comparing predicted dexamethasone sensitive and resistant ALL groups, we observed enrichment of pathways in the dexamethasone resistant group similar to that of dexamethasone-treated ALL cells including β-catenin stabilization. We also developed a drug synergy prediction model using deep learning and observed that several pathway-specific inhibitors including aurora kinase, S6K, JAK2, mTOR, etc. inhibitors displayed synergy with dexamethasone. Finally, we showed that dexamethasone displayed synergy with several β-catenin inhibitors.
Collectively, this study suggests a novel mechanism of glucocorticoid resistance in ALL in which glucocorticoids such as dexamethasone stabilized β-catenin through probably a non-canonical inactivation of GSK3β activity.6

References:
1 Wang, X., Zhang, H. & Chen, X. Drug resistance and combating drug resistance in cancer. Cancer Drug Resist 2, 141-160 (2019).
2 Boumahdi, S. & de Sauvage, F. J. The great escape: tumour cell plasticity in resistance to targeted therapy. Nat Rev Drug Discov 19, 39-56, doi:10.1038/s41573-019-0044-1 (2020).
3 Shaffer, S. M. et al. Rare cell variability and drug-induced reprogramming as a mode of cancer drug resistance. Nature 546, 431-435, doi:10.1038/nature22794 (2017).
4 Inaba, H. & Pui, C. H. Glucocorticoid use in acute lymphoblastic leukaemia. Lancet Oncol 11, 1096-1106, doi:10.1016/S1470-2045(10)70114-5 (2010).
5 Hardie, D. G. Roles of protein kinases and phosphatases in signal transduction. Symp Soc Exp Biol44, 241-255 (1990).
6 Shah, K., Ahmed, M. & Kazi, J. U. The Aurora kinase/beta-catenin axis contributes to dexamethasone resistance in leukemia. NPJ Precis Oncol 5, 13, doi:10.1038/s41698-021-00148-5 (2021).
Follow the Topic
-
npj Precision Oncology
An international, peer-reviewed journal committed to publishing cutting-edge scientific research in all aspects of precision oncology from basic science to translational applications to clinical medicine.

Related Collections
With Collections, you can get published faster and increase your visibility.
Minimal Residual Disease and Circulating Tumor DNA Dynamics in Personalized Cancer Treatment
Publishing Model: Open Access
Deadline: Mar 12, 2027
Next-Generation AI in Drug Design for Precision Oncology
Publishing Model: Open Access
Deadline: Apr 01, 2027


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in