How the vasculature controls adipose tissue wasting in cancer cachexia
Published in Cancer
Background
Cachexia is a disabling wasting condition of lean body mass which occurs in response to several diseases, most notably cancer. Loss of muscle and fat mass is accompanied by decreased quality of life, poor response to chemotherapy and high mortality. Apart from curing the primary disease (in this case cancer), there are no standardized treatments to reverse cachexia. It is widely assumed that factors secreted by the tumor, such as proinflammatory cytokines, are responsible for tissue wasting. White adipose tissue (WAT) remodeling initiates early during cachexia leading to impaired lipid metabolism, insulin resistance, macrophage infiltration, chronic inflammation and eventually fibrosis.1
Given that cachexia is a whole-body response syndrome,2 we assumed that the vasculature plays a pivotal role as a mediator and potentially also executor in the pathogenesis of cancer cachexia. The barrier-forming continuous endothelium is present in most organs and prevents direct interaction between tissue and the blood’s contents. Circulating tumor-derived factors (e.g., tumor necrosis factor α (TNFα), interleukin-6 (IL-6) or IL-1b) are assumed to be, in part, responsible for tissue wasting, cannot pass the endothelial barrier and thus cannot directly act on tissue cells (e.g., adipocytes, myocytes). Our hypothesis that the endothelium itself mediates tissue wasting was based on our previous work showing that cancer-derived factors prime the vasculature at distant sites to facilitate homing of circulating tumor cells.3 More recent work demonstrated that the endothelium, the inner lining of blood vessels, systemically senses the presence of a cancer mass and responds by producing large amounts of LRG1, an endothelial-derived protein, which facilitates several steps of the metastatic cascade.4 This concept of angiocrine (paracrine) signaling plays essential roles during development and regeneration,5 and is assumed that angiokines also influence cancer progression.6
How did we get started?
In a previous project, we detected that in several tumor entities endothelial Notch1 signaling, a critical regulator of angiogenesis and angiocrine signaling,7 becomes overactivated not only within the tumor vasculature, but also at distant organs such the lung.3 In adult transgenic mice, in which we induced a moderate but sustained Notch1 signaling overactivation within the endothelium, there was not only an increase in metastatic spreading,3 but also insulin resistance,8 and WAT wasting in tumor-free mice. This was the starting point to address whether endothelial Notch1 signaling would be involved in the pathogenesis of tissue wasting.
We chose to focus on our study on the pre-cachectic stage. At this stage, WAT is known to already undergo remodeling and patients often show signs of glucose intolerance and insulin resistance. Bioinformatic analysis showed enrichment of a Notch1 gene signature in endothelial cells derived from a pre-cachectic mice and in WAT biopsies from pre-cachectic and cachectic patients with cancer, suggesting that even at this early time point in cachexia development, the endothelium may already be engaged.
Results
Our paper shows that an endothelial-specific transgenic mouse model overexpressing active Notch1 is recapitulates various hallmarks of WAT remodeling similar to mouse cachexia models and patients with cancer.
Next, we unraveled the responsible mechanisms driving endothelial-mediated WAT wasting. During pre-cachexia, we observed Notch1 signaling overactivation in the endothelium of WAT. This led to increased endothelial expression of leukocyte adhesion factor VCAM-1 and infiltration of immune cells. Bioinformatic analysis revealed that vitamin A metabolism and retinoic acid signaling were highly enriched gene signatures. ALDH1 is a family of enzymes which catalyze retinaldehyde oxidation to retinoic acid and ALDH1A2 was identified as one the most-enriched gene WAT samples from patients with cachexia. Further research revealed that Notch directly induces ALDH1 expression, retinoic acid production and downstream gene expression such as IGFBP3.
Tumor-secreted IGFBP3 has been suggested to drive key features of cachexia,9 and mediates various important pathways in adipose tissue, including IGF-1 signaling, insulin-mediated glucose uptake and adipogenesis.10 We also identified IGFBP3 induces apoptosis in vitro in our study, and likely contributes to the observed endothelial and stromal cell apoptosis.
At this time, the question arose whether retinoic acid and IGFBP3 production by endothelial cells alone is sufficient to explain the high concentration of these factors observed in WAT samples. It turned out that other cell types (adipocytes, myeloid cells) also produced high levels of retinoic acid-producing ALDH1 and IGFBP3. We ruled out that Notch1 is overactive in these cells. However, Notch1 signaling in endothelial cells led to expression of the alarmin IL-33. IL-33 could act in a paracrine manner on adipocytes and myeloid cells to induce ALDH1 expression and retinoic acid production. As such, sustained endothelial Notch1 signaling initiates a vicious cycle resulting in excessive retinoic acid production in WAT. Pharmacological inhibition of retinoic acid signaling prevented WAT remodeling during pre-cachexia, thus supporting our hypothesis.
Translational impact
We could show that induction of endothelial Notch1 signaling and Notch1-induced ALDH1, IL33 and IGFBP3 expression leads to WAT remodeling in a pancreatic ductal adenocarcinoma mouse model, which was mitigated by pharmacological inhibition of retinoic acid signaling. This indicates that the Notch-retinoic acid-IGFBP3 signaling axis may serve as an attractive target to prevent WAT wasting during cancer cachexia.
Open questions and Outlook
An unresolved question is which tumor-derived factors cause endothelial Notch1 overactivation. Our data indicate that IL-1β and TNFα, both well-known pro-cachectic factors, lead to gene induction of the Notch ligand Jagged-1. However, additional circulating molecules might be involved in this process.
An outstanding question in the cachexia field is why males are more severely affected than females.11 In line with our study, most of the phenotypes were only observed in male but not female transgenic mice, however, the underlying mechanisms remain elusive.
Lastly, it will be of utmost importance to carefully test whether long-term pharmacologic blockade of retinoic signaling is tolerable and whether it can inhibit WAT wasting in a large set of preclinical cancer models.
Figure
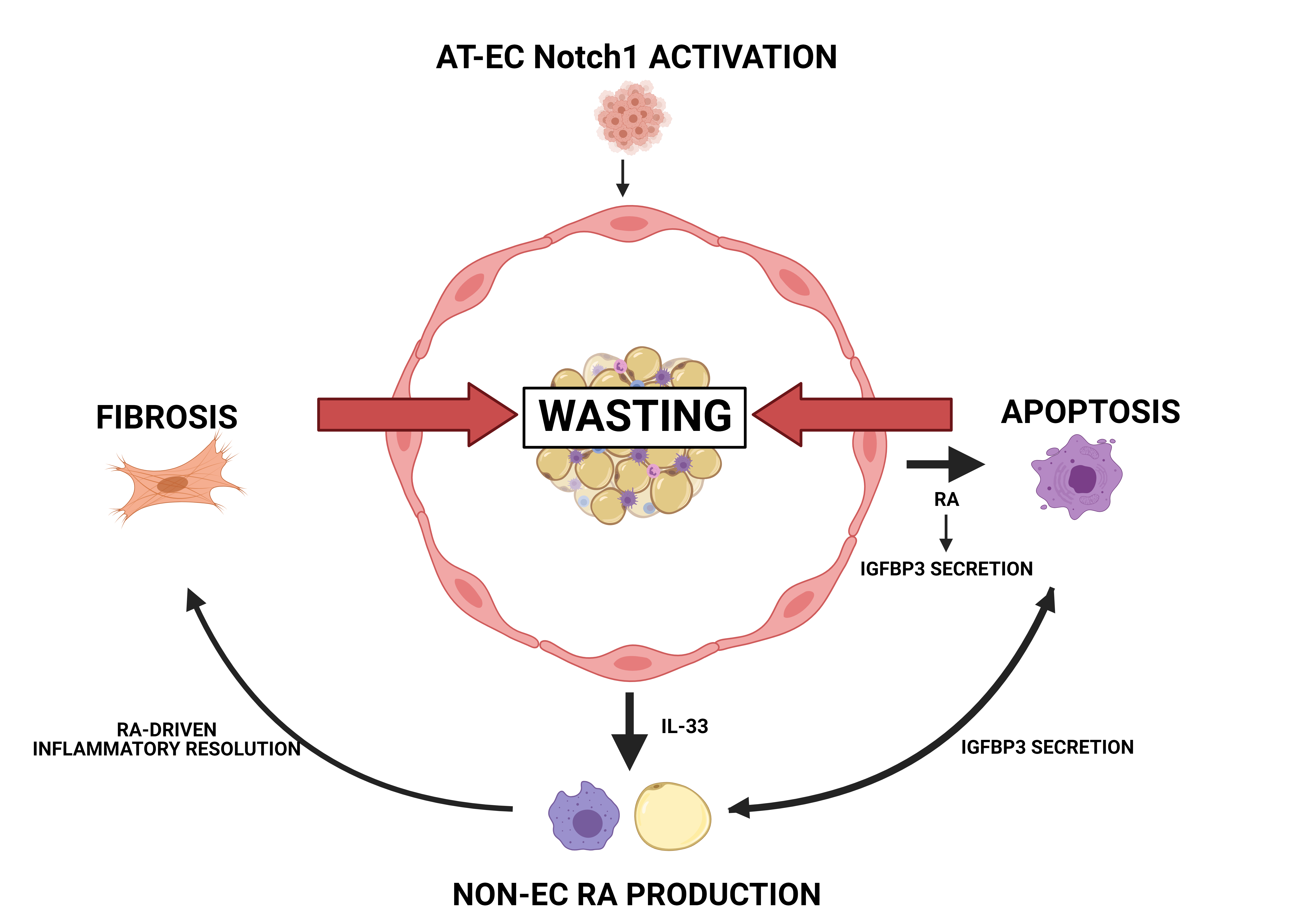
Figure legend
Tumors activate Notch1 in the endothelium (AT-EC) within WAT. This activation leads to the production of retinoid acid (RA) and the secretion of IGFBP3. In addition, Notch signaling leads to the secretion of IL-33, which, together with IGFBP3, further induces RA production in macrophages and adipocytes. Finally, RA promotes a continuous resolution of inflammation, which leads to fibrosis and eventually WAT wasting.
References
1 Schmidt, S. F., Rohm, M., Herzig, S. & Berriel Diaz, M. Cancer Cachexia: More Than Skeletal Muscle Wasting. Trends in Cancer 4, 849-860 (2018). https://doi.org:10.1016/j.trecan.2018.10.001
2 Ferrer, M. et al. Cachexia: A systemic consequence of progressive, unresolved disease. Cell 186, 1824-1845 (2023). https://doi.org:10.1016/j.cell.2023.03.028
3 Wieland, E. et al. Endothelial Notch1 Activity Facilitates Metastasis. Cancer Cell 31, 355-367 (2017). https://doi.org:10.1016/j.ccell.2017.01.007
4 Singhal, M. et al. Temporal multi-omics identifies LRG1 as a vascular niche instructor of metastasis. Sci Transl Med 13, eabe6805 (2021). https://doi.org:10.1126/scitranslmed.abe6805
5 Rafii, S., Butler, J. M. & Ding, B.-S. Angiocrine functions of organ-specific endothelial cells. Nature 529, 316-325 (2016). https://doi.org:10.1038/nature17040
6 Alsina-Sanchis, E., Mulfarth, R. & Fischer, A. Control of Tumor Progression by Angiocrine Factors. Cancers (Basel) 13 (2021). https://doi.org:10.3390/cancers13112610
7 Hasan, S. S. & Fischer, A. Notch Signaling in the Vasculature: Angiogenesis and Angiocrine Functions. Cold Spring Harb Perspect Med 13 (2023). https://doi.org:10.1101/cshperspect.a041166
8 Hasan, S. S. et al. Endothelial Notch signaling controls insulin transport in muscle. EMBO Mol Med 12, e09271 (2020). https://doi.org:10.15252/emmm.201809271
9 Martin, A. I. et al. IGF-1 and IGFBP-3 in Inflammatory Cachexia. Int J Mol Sci 22 (2021). https://doi.org:10.3390/ijms22179469
10 Baxter, R. C. & Twigg, S. M. Actions of IGF binding proteins and related proteins in adipose tissue. Trends Endocrinol Metab 20, 499-505 (2009). https://doi.org:10.1016/j.tem.2009.07.002
11 Zhong, X. & Zimmers, T. A. Sex Differences in Cancer Cachexia. Current Osteoporosis Reports 18, 646-654 (2020). https://doi.org:10.1007/s11914-020-00628-w
Follow the Topic
-
Nature Cancer
This journal aims to provide a unique forum through which the cancer community will learn about the latest, most significant cancer-related advances across the life, physical, applied and social sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Cancer Neuroscience: from mechanisms to therapy
Publishing Model: Hybrid
Deadline: Jan 30, 2027


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in