It is the bane of a microorganism’s existence to have to co-exist with multicellular organisms with complex physiologies on a planet that is growing increasingly uninhabitable. Although cursed with comparatively simple biology, these amazing creatures are extremely adaptable and have evolved complex machineries that function together allowing them to survive extreme fluctuations in availability or a dearth of environmental nutrients. Transition metals, such as Zn, are one such class of nutrients while extremely important for several vital physiological processes, like most things in life, turn toxic when their concentrations rapidly fluctuate as microorganisms encounter changing environmental conditions.
When I joined Prof. Peng Chen’s lab as a graduate student, I was handed the daunting task of trying to understand how a certain class of proteins, called metalloregulators, work together to sense and control the Zinc levels inside bacteria. From spending endless hours in a dark microscope room with flashing lasers to analyzing results during the uncertain lockdown era away from family on a different continent, this project was a test of character, and I not only discovered an unprecedented ‘through-DNA’ mechanism for metal-regulation but also discovered my love for single-molecule biophysics.
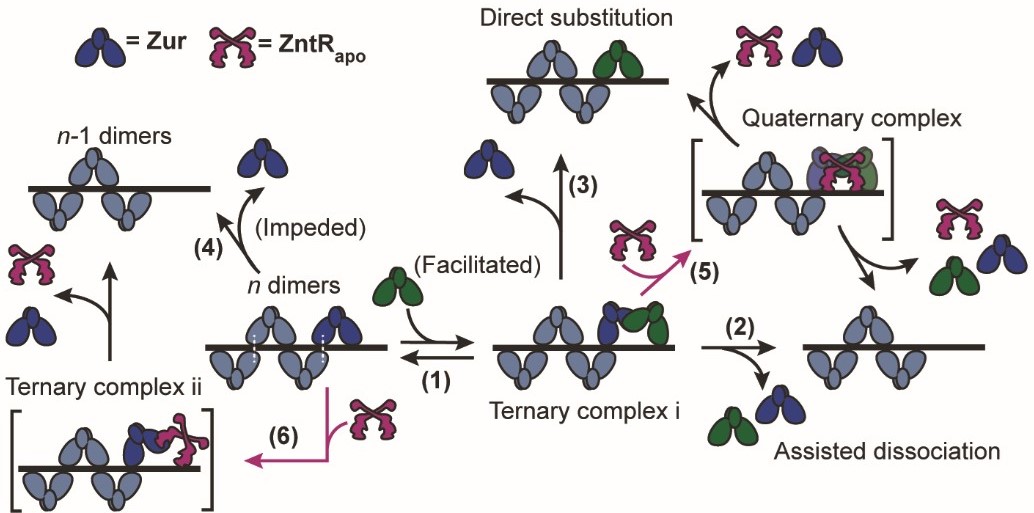
Metalloregulators, involved in sensing and regulating metal uptake and efflux, are conventionally believed to interact with each other indirectly by sensing the environmental metal concentrations, without physically interacting with one another. When bacteria face rapidly changing metal environments, in-vivo regulation status needs to quickly reset (Figure 1a). Our lab had previously discovered a ‘facilitated unbinding’ mechanism, that allows homotypic metalloregulators to unbind from DNA, quickly switching the transcription of regulon efflux and uptake proteins, on or off, depending upon the cellular metal requirements. This mechanism involves a cytoplasmic metalloregulator, that either dislodges or replaces the incumbent metalloregulator from its binding locus on the DNA, operating via the formation of a ternary complex in which both proteins bind partially to a dyad-symmetric DNA sequence (Figure 2e).
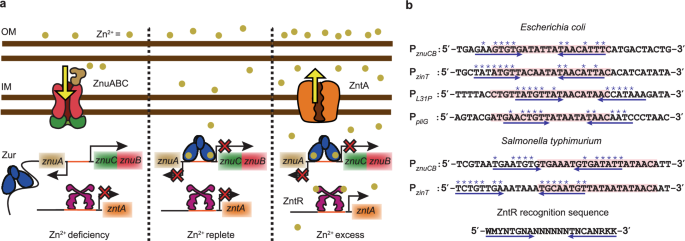
We focused our current research on a pair of prototypical Zn uptake and efflux regulators Zur and ZntR in E. coli, and upon analyzing the regions of the DNA that Zur recognizes, we surprisingly discovered overlapping sequences that bore partial similarities with the regions of DNA that ZntR recognizes (Figure 1b). This interesting arrangement of partial efflux regulator binding sequences next to where the uptake regulator binds on DNA made us wonder whether ZntR could bind next to Zur and help to enhance Zur unbind. We used a combination of in-vivo single-molecule tracking and stroboscopic imaging to track single Zur proteins that were tagged with a photo-convertible fluorescent protein, mEos3.2. We also tagged ZntR with another fluorescent protein super-folder GFP that allowed us to spectrally separate the two proteins and estimate cellular ZntR concentrations. To mimic toxic Zn environments where the proteins are fully metallated, cells were grown stressed with 20uM Zn; Further, specific genetic mutations were made on Zur (Cysteine88 → Serine) and ZntR (Cysteine115 → Serine) to mimic their apo forms that exist under conditions of metal deficiency. Individual cells were then sorted into groups having similar Zur and ZntR concentrations and compiling several Zur displacement trajectories within each concentration group allowed us to extract Zur unbinding kinetics as a function of both protein concentrations.
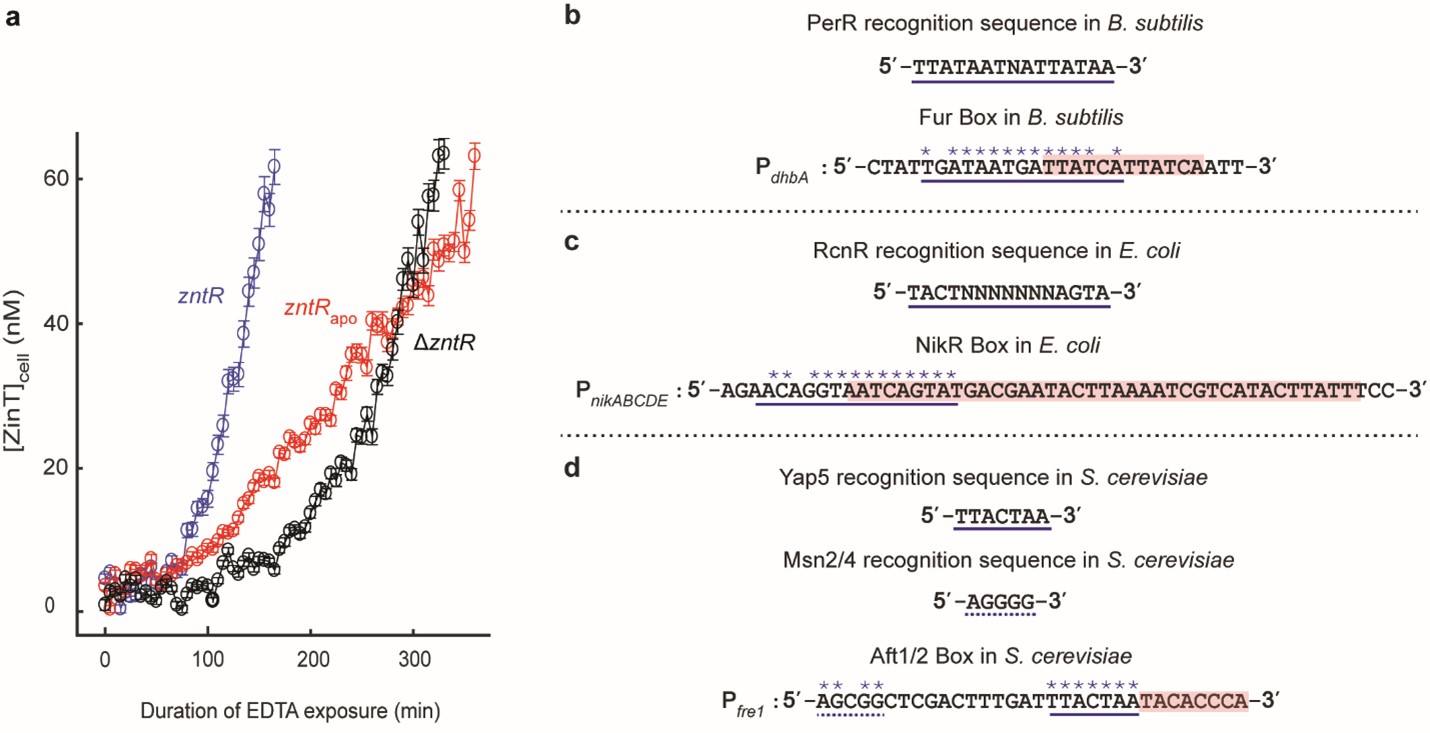
For both the Zn replete (Zn sufficiency condition; where Zur exists in its metallated form and ZntR remains in its apo form) and the deficiency conditions, we strikingly observed that as the cellular ZntRapo concentration increased, the Zur unbinding rate constant, k-1 (Figure 2a and b) also increased. While, for the conditions of Zn excess, we observed a negligible enhancement of Zur unbinding with increasing ZntRZn concentration, which seems reasonable considering ZntR primarily functions as an efflux regulator under these conditions and would prefer Zur to remain bound and repress Zn-uptake (Figure 2c). We further observed the Zur binding rate constant, k1 remains independent of ZntR concentration (Figure 2d), suggesting that ZntR does not bind to the partial DNA sequence on its own to block Zur binding on DNA and instead requires Zur to be bound on the DNA, possibly involving Zur-ZntR interactions besides the partial ZntR-DNA interactions. To ensure the directness of these effects, we purified Zur/ ZntR and labeled Zur with the fluorescent dye, Cy5 and performed in vitro single-molecule FRET measurements, with a commercially purchased DNA labeled with Cy3 encoding both the Zur and the partial ZntR binding sequence. We observed a consistent enhancement of Zur unbinding from the DNA as a function of ZntR concentration. These experiments were majorly conducted by my collaborator, Dr. Youngchan Park. Alongside discovering Zur and ZntR interactions, we also discovered a deep friendship, inspiring each other when the going got tough.
![Figure. 2 | a, Dependence of the apparent unbinding rate constant k-1 of on its own concentration and at different [] in the cell. b, Dependence of the apparent unbinding rate constant k-1 of on its own concentration and at different [] in the cell. c, Same as (a), but for k-1 of and at different [] in the cell. d, The binding rate constant k1 of vs. cellular [] or [. Solid line: linear fit; dashed line: horizontal line fit. Error bars are SEM. f, Mechanistic model for ZntRapo-dependent Zur unbinding kinetics. Starting with oligomerized Zur (dark and light blue) at a tight-binding site on DNA, the unbinding of an incumbent Zur protein (dark blue) can be facilitated by a freely diffusing Zur (dark green) through the formation of a ternary complex i (step 1), leading to assisted dissociation (step 2) or direct substitution (step 3); this facilitated unbinding of Zur can be enhanced by ZntRapo through the formation of a heteromeric quaternary complex (step 5). The oligomer-induced impedance of Zur unbinding (step 4) can be weakened by ZntRapo through the formation of a heteromeric ternary complex ii (step 6), leading to faster Zur unbinding as well. White dashed lines on the ‘n dimers’ denote salt bridge interactions between Zur dimers.](https://images.zapnito.com/cdn-cgi/image/metadata=copyright,fit=scale-down,format=auto,quality=95/https://images.zapnito.com/uploads/uhx9Jj03S22MCPQRZKiX_picture5.jpg)
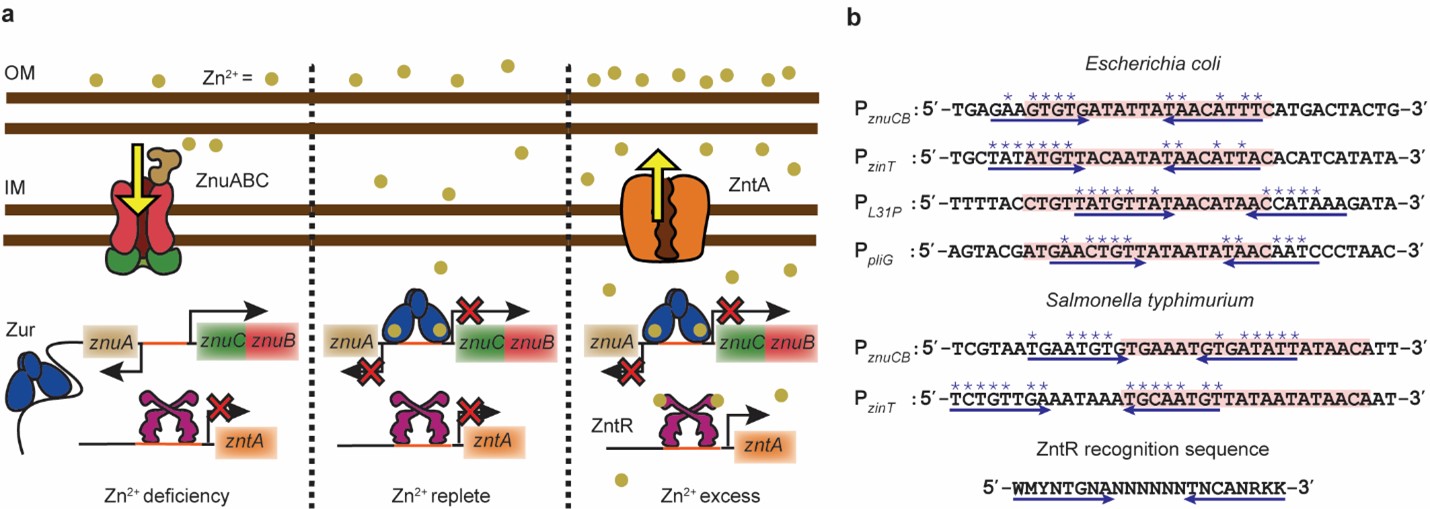
To assess the physiological relevance of this effect, we studied the derepression of a Zur, regulated protein, ZinT, tagged with super-folder GFP, as a function of EDTA mediated Zn-depletion. Compared to the strain where the zntR was knocked out, in the strain containing ZntRapo, we observed a faster derepression of ZinT (Figure 3a), suggesting a role of ZntR to facilitate Zur unbind as cells transition from conditions of Zn excess to deficiency, necessitating an induction of uptake proteins. We also analyzed uptake regulator binding sequence, for other different metal-regulator pairs, in different bacteria, and even in higher organisms like yeast (Figure 3b-d), and discovered a similar partial overlap between efflux and uptake regulator sequences, suggesting a broader relevance and a possible wider prevalence of this mechanism.
In a world where “life breaks free–[crashing] through barriers painfully, maybe even dangerously”, antibiotic resistance is a global crisis. Thus, identifying such underlying mechanistic paradigms, besides throwing additional light into metal homeostasis, could open avenues for the development of efficacious metal-based drugs and treatments that could target these pathways, curbing pathogenic infections mitigating a global threat of antibiotic resistance.
Another Blog Post by my collaborator and co-first author, Dr. Youngchan Park is available at: https://go.nature.com/4imxwgu
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in