
Our laboratory studies irritable bowel syndrome (IBS), a complex, heterogenous disorder characterized by chronic symptoms of food intolerance, abdominal pain and altered bowel habits. Although a number of studies have demonstrated microbiota alterations in IBS patients, only a handful of these studies have assessed functional significance of these changes. Despite significant interest and investment, no clear breakthrough has been made in microbiota-based therapies for IBS and current restoration strategies (probiotic or fecal microbiota transplantation, FMT) have yielded poor and mixed results1. One commonly identified subset of IBS is post-infection IBS which develops after an episode of gastroenteritis such as Campylobacter jejuni. Clinically, this has been a fascinating subset for us as these individuals are normal prior to the infection and subsequently develop chronic and often disabling GI symptomatology. Our previous findings had shown that 40% patients with post-infection IBS demonstrated increased fecal proteolytic activity (PA), lower microbial diversity and changes in microbiota composition from rest of the IBS patients or matched healthy volunteers2. In the current study, we embarked on determining the nature of proteases driving the PA and the broader regulation of PA in the gut. We envisioned this would be of relevance not only for IBS but for variety of other disorders of the gut which have been associated with aberrant regulation of PA (like ulcerative colitis, Crohn’s disease and celiac disease)3. Moreover, the intestinal proteases play a fundamental role in digestive and defense processes. Our journey in determining the regulation of PA took us through multiomics in human and mice samples as well as several, targeted intervention studies in mice.
Host and microbial proteases can both drive fecal PA, however, the net contribution of these towards the overall fecal PA has remained elusive. We first tasked ourselves with determining host and microbial peptides in human stool using untargeted proteomics strategies. Utilizing tools developed by the Galaxy pipeline (http://galaxyp.org/), we determined three proteins to be differentially abundant between high and low PA samples, all serine proteases of human origin (Trypsin-2, Chymotrypsin-like pancreatic elastase 2A and 3B). Microbial proteases were identified but these were not significantly different. The colonic epithelial RNA or protein expression was not different, suggesting these proteases are likely of pancreatic origin. These findings left lingering, important questions regarding the mechanisms that regulate luminal PA. Microbiota have been well documented to have various mechanisms which inhibit or support the activity of proteases for both defensive and competitive advantages in local niches and their environment. As a result, we investigated intestinal microbiota and its role in PA regulation. We started with simplified approaches using germ-free mice. We were intrigued to observe that germ-free mice had significantly higher fecal PA than their conventionally raised counterparts (over 10-fold). This was an exciting observation, and for us, it raised a new question: Does introducing microbiota to germ-free mice result in changes in PA, and more importantly, will this differ with specific human microbial communities?
We humanized germ-free mice, a process where we introduce specific intestinal microbiota to mice using fecal samples taken from volunteers via oral gavage. The engrafted microbiota colonizing the GI tract represents a composition which can serve as a surrogate to study the human donor’s microbiome. Upon humanization we found that healthy commensal microbiota suppressed the PA significantly, but this effect was not seen after humanization using microbiota from high PA IBS patients (75% vs 0% inhibition respectively). Moreover, we observed mice humanized with high PA microbiota had increased in vivo intestinal permeability compared to the healthy humanized group. These observations indicated that microbial communities within the GI tract have a significant role in regulating fecal PA and gut function.
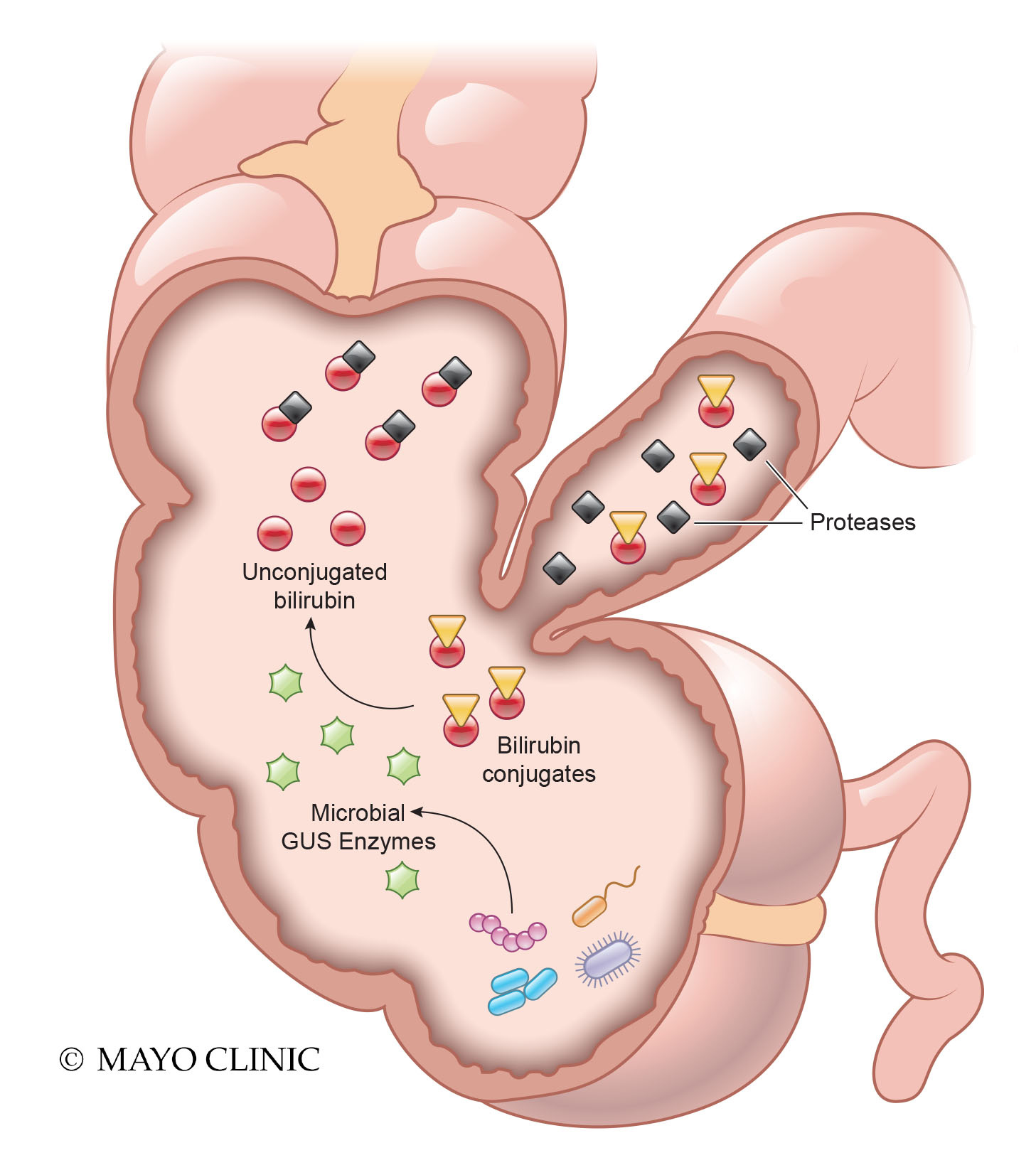
Next, we wanted to identify the mechanism by which microbiota suppress host PA. Our analysis of high PA communities found key bacterial species were missing compared to healthy communities. Machine-learning strategies demonstrated Alistipes putredinis to be a leading missing microbial taxon in high PA samples in both patients and mice. Upon supplementation of these bacteria through fecal microbiota transplantation, PA was suppressed. Interestingly, the missing bacteria from high PA communities produce an enzyme β-glucuronidase (GUS). Microbial derived GUS enzymes play an important role in deconjugation of bilirubin glucuronides produced by the liver and to use glucuronic acid as a carbon source. High PA IBS patients had lower fecal GUS activity and end-products of bilirubin metabolism Our study showed following key experimental findings in mice that demonstrate a microbial GUS driven process of PA regulation 1) high PA mice gavaged with unconjugated bilirubin dropped their PA, 2) high PA mice administered GUS overexpressing E. coli dropped their PA, and 3) GUS inhibitors resulted in increase in PA in high PA mice. Taken together, our studies in humanized mice establish an important role for intestinal microbiota in inhibiting host proteases through the production of unconjugated bilirubin via microbial GUS activity.
These studies taught us the power of hypothesis-driven multiomics studies to answer questions raised by observation in patients and then using microbiota-dependent mice models to mechanistically understand the biological process. These findings set us up for questions for the future in understanding precise GUS machinery and their microbial sources that are most effective in deconjugating bilirubin and ultimately proteolytic activity suppression. Furthermore, translating these findings for providing targeted microbiota-based therapies for IBS would be attractive. As a patient- and disease-oriented laboratory, we are excited for the next frontier of research and clinical trials in mechanism-driven microbiota-based targeting for alleviating IBS symptoms.
1. Xu, D. et al. Efficacy of Fecal Microbiota Transplantation in Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis. Am J Gastroenterol 114, 1043-1050, doi:10.14309/ajg.0000000000000198 (2019).
2. Edogawa, S. et al. Serine proteases as luminal mediators of intestinal barrier dysfunction and symptom severity in IBS. Gut, doi:10.1136/gutjnl-2018-317416 (2019).
3. Vergnolle, N. Protease inhibition as new therapeutic strategy for GI diseases. Gut 65, 1215-1224, doi:10.1136/gutjnl-2015-309147 (2016).
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in