MacroH2A histone variants modulate enhancer activity to repress oncogenic programs and cellular reprogramming
Published in Cancer
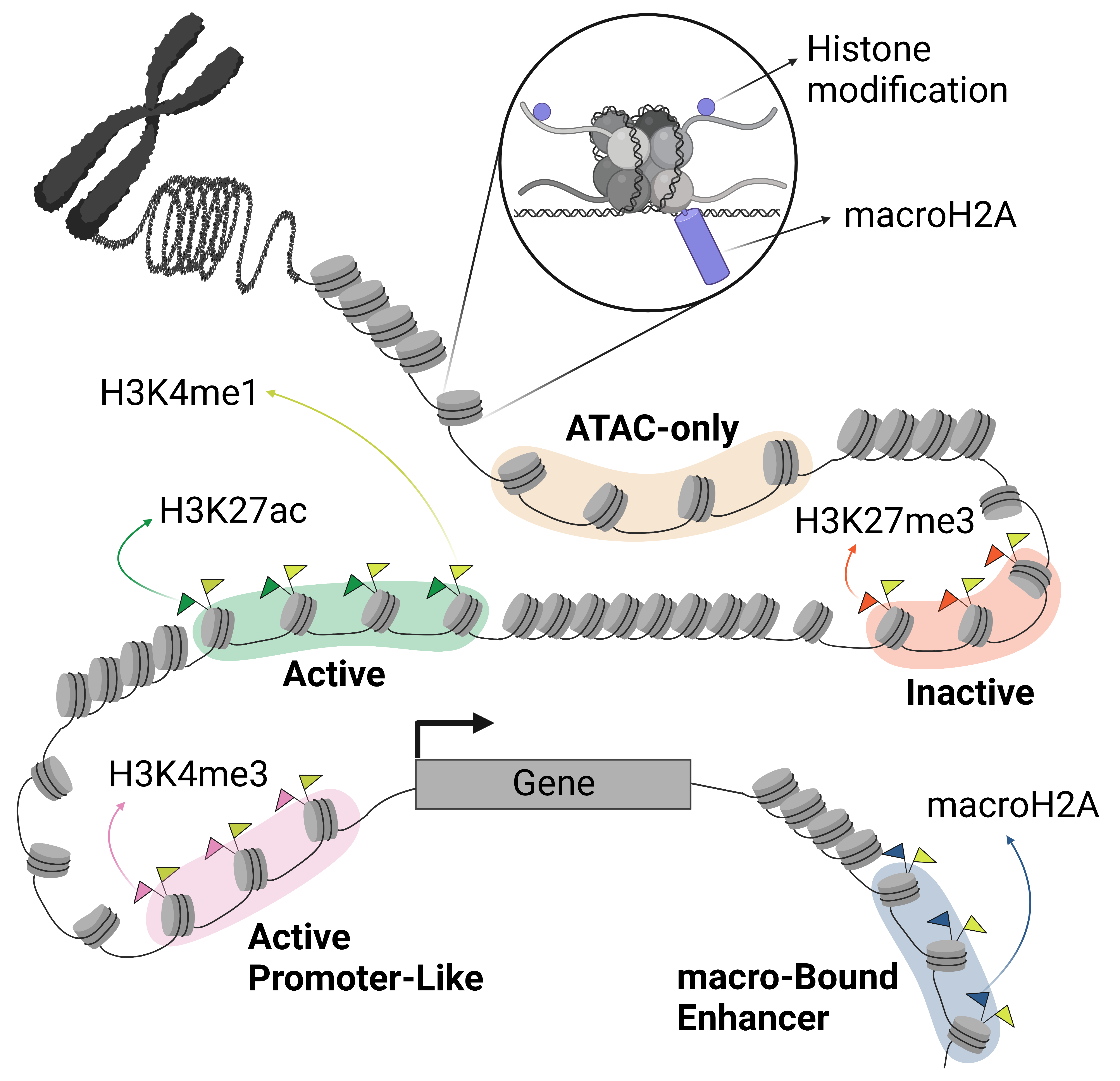

Enhancers are regions in the DNA that are bound by transcription factors (TFs) which regulate gene expression. These regions are known to be flanked by histones harboring specific modifications. Combinations of these histone modifications have been broadly utilized to annotate functional regions of chromatin1. Like histone modifications, histone variants are also known to mark specific regions of the chromatin. MacroH2A (mH2A) histone variants have been shown to be associated with various forms of inactive chromatin states. They have also been implicated in the maintenance of cell identity acting as a barrier to somatic cell reprogramming. In addition, it has been shown that the expression of specific mH2A isoforms is reduced in several cancer cell types2. It is important to note the biological parallels between reprogramming and oncogenesis – both processes involve the transformation of differentiated cells into a more undifferentiated states and are characterized by global epigenetic changes.
Since mH2A typically covers large chromatin domains we sought to understand a potential role for these histone variants in regulating enhancers. We hypothesized that the presence of mH2A in enhancer regions may play an important role in maintenance of cell identity and oncogenesis. To validate this hypothesis, we first looked at the global distribution of mH2A along the chromatin in three cell types and found that their presence on average is higher in enhancer regions and some inactive regions, compared to all other regions. But their pattern of distribution across different enhancer states was not uniform and differed by cell type. This led us to believe that a subset of enhancers was bound by the mH2A depending on the cell type. To characterize this subset of enhancers in each cell type, we designed a k-means clustering based computational method. This algorithm utilized data from several epigenetic assays to identify cell type specific regulatory elements and classify them into five functional groups including one group of enhancers enriched in mH2A which we termed macro-Bound Enhancers (mBE).

But what do mBEs do?
To probe the functional role of mBEs, we performed cellular reprogramming in cells derived from mH2A double knockout (dKO) mice. In reprogramming experiments, adding a special cocktail of TFs - Oct4, Klf4, Sox2 and cMyc (Yamanaka factors), triggers the transformation of differentiated (somatic) cells into induced pluripotent stem (iPS) cells. Since mH2A variants have been previously shown to act as a barrier to reprogramming3, we hypothesized and subsequently validated that mBEs are enriched at the binding sites of these factors, most significantly at the enhancers of the pioneering factors Oct4 and Sox2. Using a CRISPR/Cas9-mediated epigenome editing system to bring the macro domain (the non-histone domain of mH2A) to targeted enhancer regions, we showed that the macro domain inhibits gene expression similar to other known repressor domains such as the KRAB domain. Thus, the presence of mH2A at enhancers during reprogramming may hinder their activation and inhibit reprogramming.

Do mBEs have any role in cancer?
Since mBEs potentially serve as gatekeepers of cell identity, we hypothesized that the loss of mH2A in cancer could lead to oncogenic gene expression programs by means of enhancer activation. Analysis of epigenetic data from breast cancer subtypes suggested that mBEs function in a context dependent manner and are associated with breast cancer risk variants from genome-wide association studies. Because mH2A2 variant had the strongest repressive effect in the reprogramming assays, we modeled the role of mH2A2 using breast cancer cell lines that represent two extremes in terms of mH2A2 expression - MCF7 (high) and MDA-MB-231L (low). We depleted mH2A2 in MCF7 cells by CRISPR/Cas9 genome editing, and over-expressed mH2A2 in MDA-MB-231L. Over-expression of mH2A2 led to decreased proliferative capacity.
To understand this context-dependent nature of mH2A regulation at enhancers, we defined its potential role in MCF7 cells, which is an estrogen (ER)-responsive breast cancer cell line with a well-defined enhancer network. Analysis of MCF7 specific mBEs showed preferential mBE binding of TFs associated with ER activation. Based on this observation, we hypothesized that mBEs could maintain ER-responsive enhancer elements inactive and in turn serve as a gatekeeper of the MCF7 enhancer network. So, disruption of mBE with the loss of mH2A2 should render the ER-dependent transcriptional program even more accentuated. We validated this hypothesis using a tumorsphere assay showing increased response of mH2A2 knockout (KO) MCF7 cells to estrogen when compared to wild type (WT).

Furthermore, using single cell (sc) ATAC-seq experiments, we show that there is increased interactions between mBEs and active promoter-like (APL) elements, increased activity of ER activation TFs and stem-like signatures in mH2A2 KO MCF7 cells compared to WT.

How does mBE regulation work?
To gain a better understanding of the global effect of the mH2A on enhancer regulation we performed epigenetic profiling of MDA-MB-231L cell model. We found that gain of function of mH2A2 led to loss of chromatin bound BRD4, an epigenetic reader that binds active enhancers and promoters. BRD4 was also mostly excluded in mBE in WT MCF7 cells. This suggests a potential mechanism by which mBEs render enhancers inactive.

Next, we investigated the potential role of mH2A in restraining mammary epithelial cells from oncogenic programs. By investigating mammary gland development in wild type and mH2A dKO mice, we showed that in some mH2A dKO mice the mammary epithelial cells present a more stem-like state. Using single cell multiome assay in organoids grown from mouse mammary epithelial cells, we showed that the mH2A dKO organoids have increased proportion of progenitor (stem-like) cells with increased stem cell signatures in both gene expression and open-chromatin profiles, and increased Brd4 binding sites. Evidence from our data suggests that mH2A deficiency in the mammary gland contributes to epigenetic plasticity, which could potentially lead to activation of oncogenic programs.

In summary, we characterized a specific class of enhancers that are marked by enrichment of macroH2A histone variants and showed that these enhancers play a role in preserving cell identity through cell-specific modulation of transcription, with important implications for cellular reprogramming and activation of oncogenic pathways.
References
- The ENCODE Project Consortium., Moore, J.E., Purcaro, M.J. et al. Expanded encyclopaedias of DNA elements in the human and mouse genomes. Nature 583, 699–710 (2020).
- Ghiraldini, F.G., Filipescu, D. & Bernstein, E. Solid tumours hijack the histone variant network. Nat Rev Cancer 21, 257–275 (2021).
- Gaspar-Maia, A., Qadeer, Z., Hasson, D. et al. MacroH2A histone variants act as a barrier upon reprogramming towards pluripotency. Nat Commun 4, 1565 (2013).
Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

Related Collections
With Collections, you can get published faster and increase your visibility.
Artificial Intelligence Methodology in Structural Biology
Publishing Model: Hybrid
Deadline: Nov 30, 2026
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in