Nuclear ANLN regulates transcription initiation related Pol II clustering and target gene expression
Published in Cancer and Cell & Molecular Biology
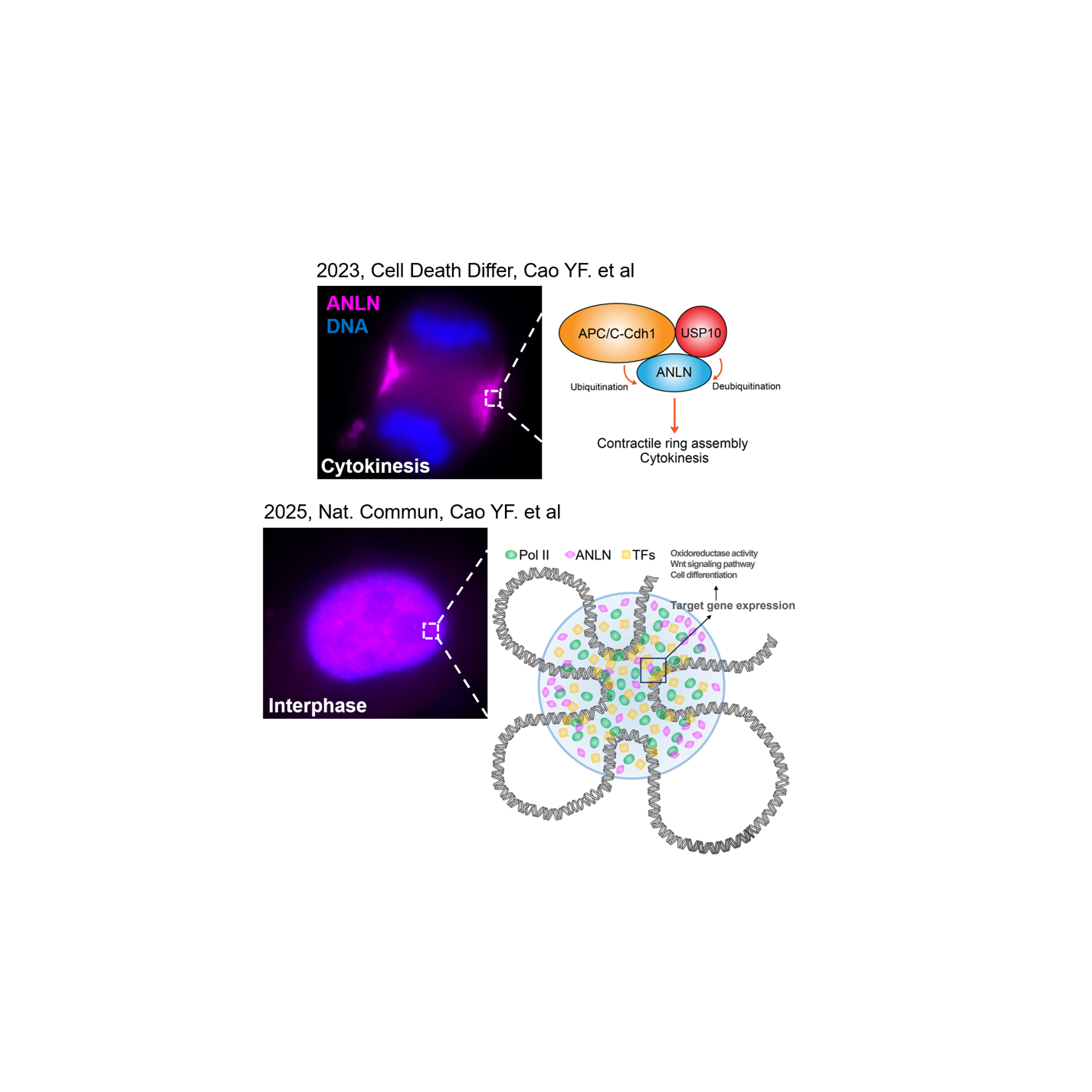

The development of malignant tumors is based on the rapid division and proliferation of cancer cells. Due to the abnormal expression and modification of mitotic proteins, cell division of cancer cells is usually uncontrolled. Anillin (ANLN) is a mitotic protein that promotes mitosis by organizing contractile ring components, a process that lasts approximately 40-60 minutes. ANLN is overexpressed in a variety of cancers, promoting cell proliferation, and is closely related to patient survival. In particular, our group previously found in esophageal squamous cell carcinoma (ESCC) that the protein level of ANLN is controlled by ubiquitination and deubiquitination, thereby regulating the division and proliferation of ESCC cells (DOI: 10.1038/s41418-022-01104-x). However, during an interphase of up to 20 hours, ANLN is distributed in the nucleus and may play unknown functions. It is difficult to fully explain the cancer promoting mechanism of ANLN only by regulating contractile ring assembly. To this end, we carried out a two-year research exploration to investigate the role of nuclear ANLN in cancer cells.
Discovery of interaction between nuclear ANLN and RNA polymerase II

Fig.1 (A) Immunofluorescence images of ANLN and DNA in cancer cells. (B) Western blotting was performed to examine the different subcellular fractions of the indicated cells. (C) Nascent RNA was labeled with BrUTP, and ANLN was labeled with antibody. (D-E) CO-IP and PLA detected the interaction between ANLN and RNA polymerase II.
We initially found that nuclear ANLN is associated with chromatin (Fig.1A, B) and nascent RNA (Fig.1C) in several cancer cells, suggesting that ANLN may play a role in gene transcription regulation. Next, the interaction between ANLN and RNA polymerase II (Pol II) was identified by protein mass spectrometry (not shown), CO-IP (Fig.1D), pulldown (not shown) and PLA experiments (Fig.1E). These findings stimulated our interest and led us to further clarify the relationship between ANLN and Pol II.

Fig.2 (A) CUT&Tag signals of ANLN and Pol II on the genome of ESCC cells. (B) Relative enrichment of CUT&Tag signals of ANLN and Pol II on TSS-gene body region. (C) Super-resolution images of nuclear ANLN and Pol II-pS5. Cells were treated with or without 1,6-HD. The number of protein clusters was quantified. (D) Raw and reconstructed images of the interaction patterns between ANLN and Pol II-pS5 large clusters.
As expected, by using CUT&Tag analysis, ANLN and Pol II were observed to colocalize on the genome, especially at transcription start sites (TSS) (Fig.2A, B). By using super-resolution imaging, it was found that ANLN and initiated Pol II formed clusters in the nucleus, which could be dissolved by 1.6-HD (Fig.2C), indicating that these clusters may be related to phase separation. Interestingly, ANLN clusters were generally enriched in the periphery of Pol II large clusters (Fig.2D), which made us wonder whether ANLN played a role in Pol II clustering.
Nuclear ANLN promotes RNA polymerase II clustering and activity

Fig.3 (A) Super-resolution images showing Pol II-pS5 clusters in ANLN rapid depletion ESCC cells introduced with ANLN WT or NLS mutant. (B) Representative images and statistical data of in vitro droplet assays with CTD-mEGFP and different concentrations of ANLN-mCherry.
Transcription factors and coactivators have been reported to contribute to the formation of initiated Pol II clusters. To explore the function of nuclear ANLN and exclude the interference of mitotic defects, we generated a Dox induced rapid knockdown cell and then introduced ANLN WT or NLS mutant. It was observed that Dox treatment for 24 h decreased the level of ANLN and reduced Pol II large clusters (type III), without affecting the small-size clusters (type I and II) (Fig.3A). The reduction of Pol II-pS5 clusters caused by ANLN rapid depletion could be rescued by ANLN WT but not NLS mutant (Fig.3A). Consistently, droplet assay showed that ANLN promoted Pol II CTD phase separation in a dose-dependent manner (Fig.3B). Interestingly, we also identified three short sequences in the disordered region of ANLN protein that mediate its interaction with Pol II, and deletion of these residues resulted in the loss of function of ANLN in promoting Pol II clustering (not shown).

Fig.4 (A) Occupancy of H3K27ac and Pol II measured by CUT&Tag in NC and ANLN depletion cells. (B) Western blot showing the whole cell lysate, chromatin pellets, and chromatin supernatant in ANLN depletion cells. (C) Pol II activity under different conditions was detected by dual-luciferase reporter assay.
Surprisingly, after short-term knockdown of ANLN, the H3K27ac signal decreased near the TSS, while the deposition of Pol II on chromatin increased (Fig.4A). This is similar to the abnormal pausing and accumulation of Pol II caused by topoisomerases or BRD4 deletion. Cellular fractionation and dual-luciferase reporter assay showed that short-term knockdown of ANLN resulted in decreased activity of Pol II (Fig.4B, C).
Identification of target genes of ANLN-RNA polymerase II axis

Fig.5 (A) Flowchart of the process used to identify the genes targeted by ANLN. (B) Relative mRNA levels of target genes were measured by RT-qPCR. (C) Representative tracks of super-enhancer regions occupancy of ANLN and Pol II. (D) A hypothetical model: ANLN, Pol II and transcription factors form an active transcriptional cluster at super-enhancer regions.
To further clarify the functional relevance of the role played by ANLN in transcriptional regulation, we identified the target genes of the ANLN-Pol II axis in ESCC cells. 84 target genes were identified by RNA-Seq combined with CUT&Tag, and they were found to be involved in cancer related pathways (Fig.5A, B). Representative targets were validated by ChIP-qPCR (not shown) and RT-qPCR (Fig.5B). Moreover, we found that ANLN knockdown resulted in decreased super-enhancer signal in ESCC cells (not shown). Consistently, the occupancy of ANLN and Pol II was found in the super-enhancer region of target genes (Fig.5C, D). Finally, we used THZ1, a super enhancer inhibitor, to inhibit the ANLN-Pol II axis and found that it effectively suppressed ESCC cell proliferation (not shown).
Our work established a direct relationship between nuclear ANLN and Pol II transcription factory, unveiling the authentic role of ANLN in transcriptional regulation, and provided a crucial insight for cancer research targeting ANLN.
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in