Pushing and pulling align forces in the ear
Published in Neuroscience, Cell & Molecular Biology, and Mathematics

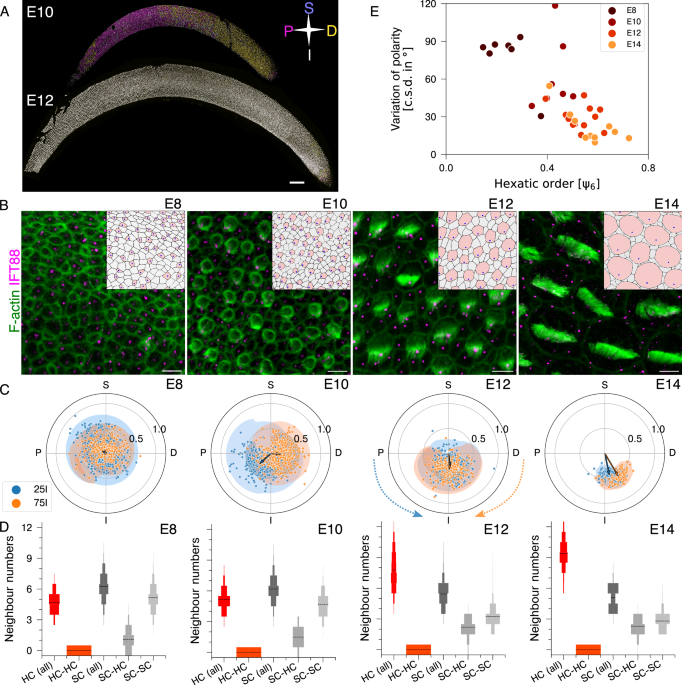
It started on a Saturday evening in the Ear Lab at NCBS, in Bengaluru, when Raj, Nishant, and I admired the patterns in the avian auditory epithelium—the Basilar Papilla (BP). The epithelium contains two key cell types: mechanosensory hair cells (HCs) and supporting cells (SCs). HCs have protrusions on their apical surfaces that move in concert to decode sound. As we observed, we realised the organisation of the BP is twofold: a spatial organisation—how cells are arranged relative to one another—and an orientational organisation—how the protrusions of each HC are aligned with those of neighbouring HCs across the epithelium. This simple appreciation sparked a bigger question that kept us busy for more than a few Saturdays: how does this organisation emerge during development?
Early on, when the HCs and SCs are first decided, HCs are smaller and surrounded by SC. The protrusions on its apical surface have just begun and the leader of those protrusions, the kinocilia, is in the middle of the apical surface. In the next two days, the kinocilia move to one edge of the cells. To our surprise, this initial movement to an edge was locally coordinated such that HCs on the one side (proximal) of the tissue had kinocilia on the proximal edge while the other side of the BP (distal) had kinocilia on the proximal edge. Over the next two days, these protrusions aligned across the epithelium in a single, unified direction, optimising sensitivity to sound. During this four-day developmental window, other dramatic changes occurred: HCs grew larger, and the BP itself elongated to twice its original length. However, even amidst this cellular turmoil, both HCs and SCs maintained a pattern where HCs were always surrounded by SCs. It was during this dynamic transformation, the twofold organisation of the BP—spatial and orientational—emerged in concert (Figure 1). 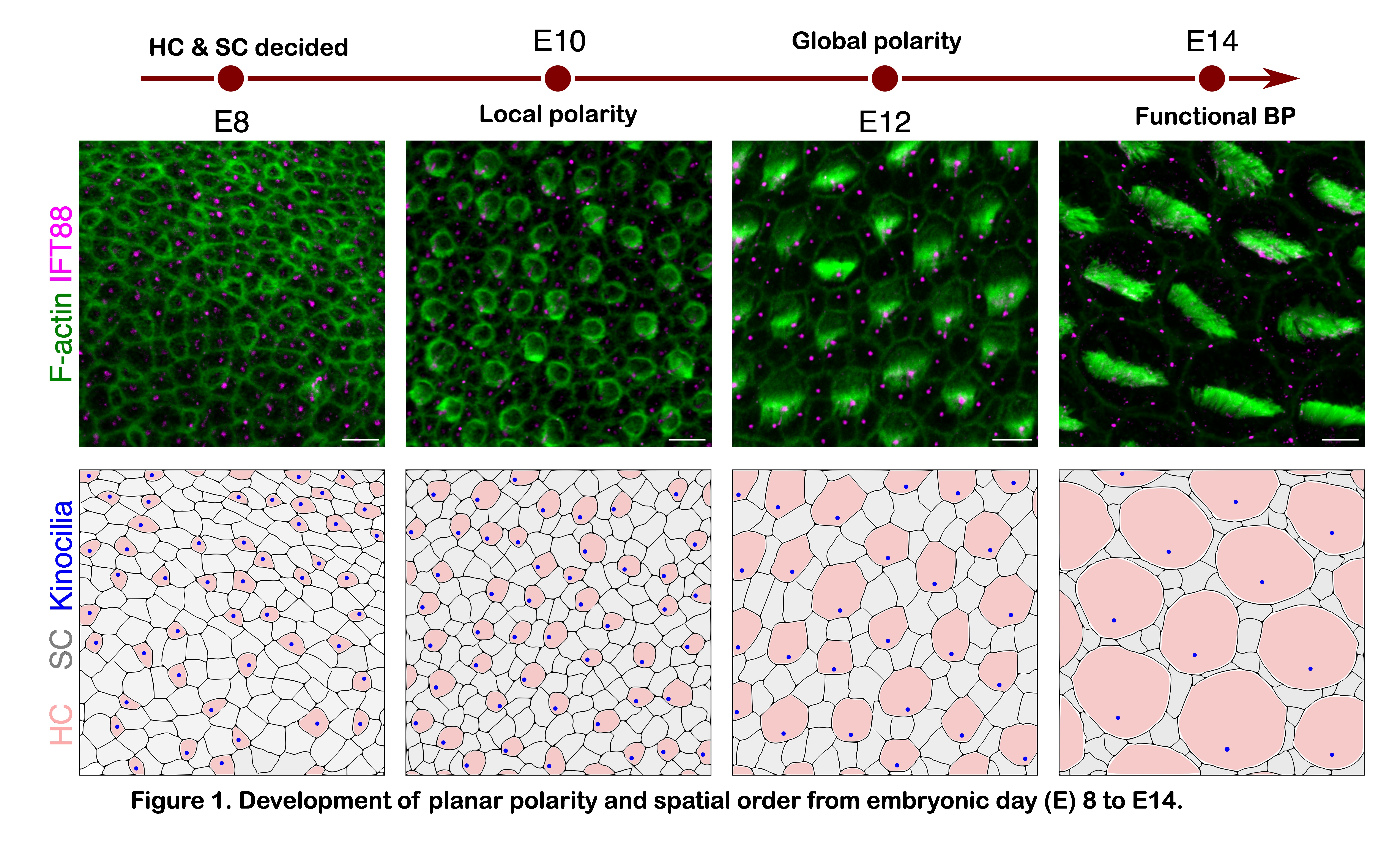
The simultaneous development of spatial and orientational order hinted at a shared underlying mechanism. We grew confident of this link when we disrupted spatial order and also disrupted orientational order. But how to find this common thread? It was intuitive to think in terms of cell neighbours—according to Lewis’ Law; larger cells tend to have more neighbours. However, in the BP, HCs and SCs with the equal surface area had different neighbour counts. This suggested a mechanical difference. Indeed, we discovered that the interface between two SCs was more contractile than between an HC and an SC. These differences were driven by the enrichment of a double-phosphorylated form of non-muscle myosin II (NMII)—essential for maintaining the HC-surrounded-by-SC architecture and, intriguingly, critical for orientational alignment too.
At that time, I was a graduate student fortunate to have Prof. Rao on my thesis advisory committee. He connected us with Prof. Karsten and Julian in Geneva. Though Julian and I were thousands of miles apart and trained in different disciplines—biology and physics—we bridged that gap through countless video calls during the pandemic. Together, we developed a physical model of our biological observations and asked,
Whether the asymmetric contractility of junctions could drive orientational ordering?
After testing multiple possibilities, we identified that the asymmetric localisation of the protein Vangl2 on SC-SC interfaces regulates the double-phosphorylated NMII, generating an asymmetric tissue axis. In HCs, the same molecular cues (LGN-Gαi) that guide kinocilium movement also shape asymmetries between the HC-SC interface near and far from the kinocilium. These two asymmetries—one in SCs and one in HCs—drive the concurrent emergence of spatial and orientational order in the BP (Figure 2).
This work marked a new beginning for us. We now see complex tissue patterns not just as shapes but as the result of intertwined spatial and orientational organisation. Such organisation, critical to physiological function, is rooted in the mechano-chemical properties of cells and the emergent behaviours arising from their interactions. This perspective allows us to think about development and organogenesis in a new way, both in vivo and in vitro.
Follow the Topic
-
Nature Communications

An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.
Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Advances in neurodegenerative diseases
Publishing Model: Hybrid
Deadline: Mar 24, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in