Shape matters and matters of shape: the interplay between cell morphology and adaptation to new environments
Published in Microbiology
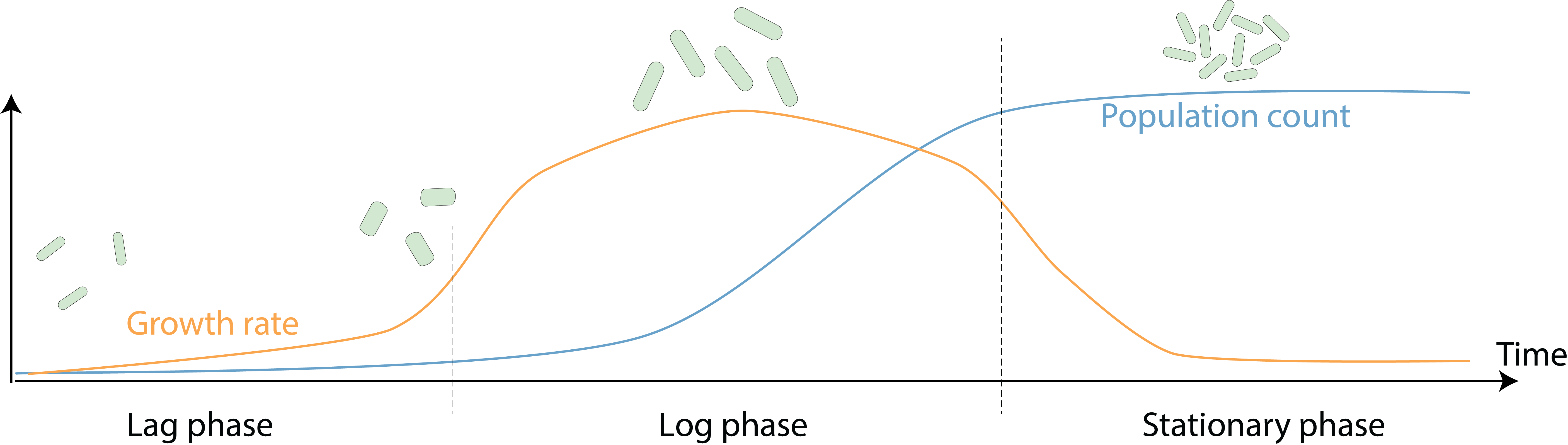
While all bacterial species have a characteristic shape and size that distinguish them from others (1), each cell also alters its cellular morphology during growth and proliferation. In the rod-shaped bacteria Escherichia coli, during a typical growth cycle in a laboratory batch culture with rich growth medium, the population number in the culture follows a sigmoidal curve: cells first gradually acclimate to the fresh nutrients and increase their growth rate, then as nutrients are depleted and waste products accumulate, growth rate slows down and population size saturates (Figure 1). Interestingly, during this process cell size is also dynamic, with wider and longer cells during fast growth, and smaller cells at slower growth rates (2, 3). Why do E. coli cells alter their morphology across growth phases? And how are morphological changes coupled to growth dynamics and external nutrient conditions? In our recent work (https://www.nature.com/articles/s41467-021-22092-5), we acquired precise and frequent single-cell measurements of cellular dimensions and growth rates from batch cultures, and developed a model that quantitatively predicts the morphological dynamics of a broad range of microbial species, highlighting the ability of cells to rapidly respond to changing environments by modifying protein synthesis and physical growth.
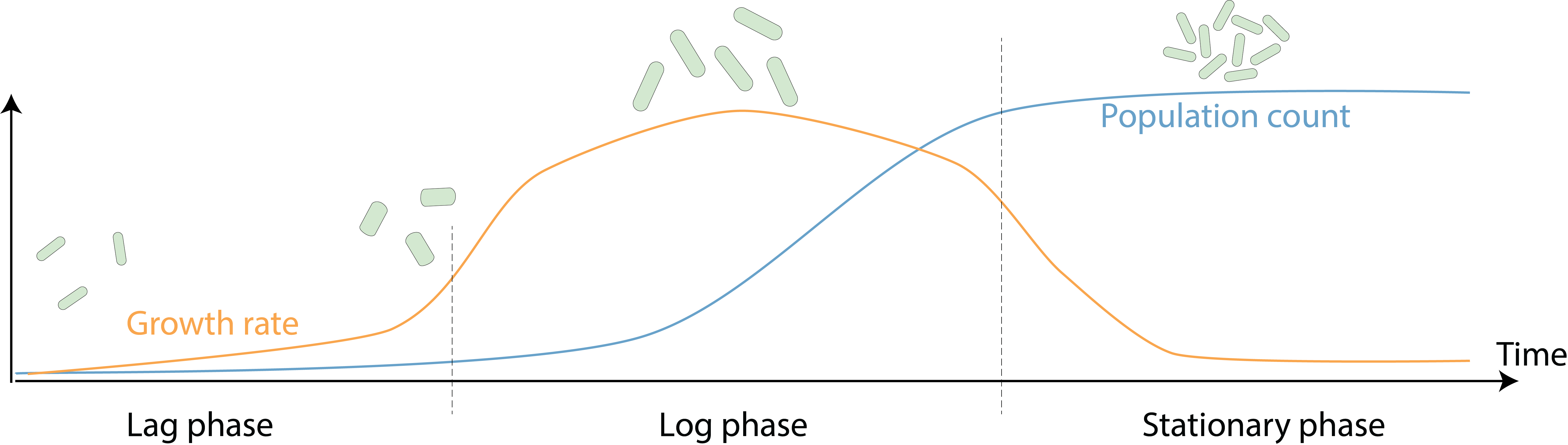
Figure 1. A typical growth curve in a laboratory culture. When saturated cell cultures are diluted into fresh medium, they first gradually increase their growth rate during lag phase. Then, cells enter log phase, in which growth rate reaches its peak and cells quickly replicate until nutrients are depleted. Cells eventually enter stationary phase, with slowed down growth and largely constant population size. During this process, cells also actively tune their morphology: exit from lag phase is first accompanied by an increase in cell width, then cell length starts to increase and allows cells to divide in log phase. Eventually, cells reduce both length and width again when entering stationary phase.
This project was motivated by our previous studies, in which we sampled cells from a batch E. coli culture at different time points, and found that changes in cellular dimensions are not isotropic (2, 3) – in other words, while cell width and length are both higher in log phase, cells do not simply scale their width and length in the same way. In fact, the increases in width and length are also not synchronized. Minutes after cells are diluted into fresh medium, they start to increase in width, but the increase in length does not occur until approximately 30 minutes later (Figure 1). This observation prompted us to consider that growth by increasing width versus length may have specific advantages to cells.
An obvious difference between increasing cell width instead of length is that an increase in width leads to a lower surface area to volume ratio (SA/V). Therefore, when cells increase their volume by the same amount, they economize on surface area expansion when growing in the width direction as opposed to length. Indeed, during growth in a batch culture, cells first decrease SA/V as they increase growth rate. Afterward, when nutrients have been depleted and growth slows down, SA/V gradually reverts back to the initial levels. Indeed, a previous study discovered a simple model of SA/V regulation at steady state when cells are shifted between two nutrient conditions supporting different growth rates (4). Thus, we attempted to use this steady-state SA/V model to explain the observed SA/V dynamics in our batch cultures, in which growth rates constantly change in response to nutrient consumption. However, the steady-state model failed (unsurprisingly) to predict the dynamics in batch cultures. Through careful inspection, we found that the steady-state model fails to capture a key factor in dynamic environments – the synthesis rates of surface area and volume do not change at the same time. In fact, using multiple measurements, including proteomics and single-cell fluorescence microscopy, we found that the changes in SA-related proteins always exhibited a 10-minute delay compared to changes in volume-related proteins. After incorporating the time delay as a fitting parameter into the model, we were able to achieve a near-perfect fit to the experimental data.
We further extended our measurements to other rod-shaped organisms, including other bacteria species and even budding yeast, a eukaryotic fungus. In all cases, we observed similar SA/V dynamics that were well-fit by our time delay model. We were also able to systematically perturb SA/V dynamics: our time-delay model predicts that when SA synthesis is limited without changing volumetric growth rate, SA/V drops. Also, with a longer time delay between SA and volume synthesis, log-phase SA/V is predicted to decrease while stationary-phase SA/V increases. We were able to experimentally verify both predictions using antibiotics targeting cell wall synthesis and protein translation, respectively. Such successes in predicting SA/V dynamics further highlight the biological relevance and utility of our time-delay model.
Finally, coming back to the original question – why do cells increase their width when exiting stationary phase but then decrease again? Our model shows that the initial increase in width allows SA/V to drop more quickly, meaning that the cell is prioritizing volumetric growth over surface area growth. However, eventually cells need to divide, and division adds two hemispherical poles to the cell body, increasing surface area. The surface area of the additional poles increases with cell width, and therefore a wider cell would need more materials to build the poles, which potentially limits the width of cells. Indeed, we observed that after exiting stationary phase, cell width starts to decrease before the first division occurs. The timing of the first division also correlates with the increased expression of a key division protein, FtsZ. FtsZ levels remain low for quite a long time after dilution to fresh medium, and its upregulation is even slower compared to other surface area synthesis proteins, presumably because the need for division comes even later after inoculation into fresh medium. Therefore, upon exposure to fresh nutrients, cells first prioritize their resources on volumetric growth, then surface area growth, and eventually cell division. Such temporal dynamics of protein expression further reinforce the concept that cells actively regulate protein synthesis as needed to maximize resource utilization.
Taken together, our work reveals a resource allocation strategy for bacterial cells under dynamic environments. The time-delay model enables quantitatively accurate predictions of cell size and shape under different growth conditions and is generally applicable to single-cell microbes. In eukaryotic cells, a nuclear transporter receptor directly senses SA/V (5). Moving forward, it remains to be discovered whether similar sensors for SA/V exist in bacterial cells, which will provide the potential to reveal fundamental connections between cell growth, physiology, and morphogenesis.
References
- K. D. Young, The selective value of bacterial shape. Microbiol. Mol. Biol. Rev. 70, 660-703 (2006).
- H. Shi et al., Deep phenotypic mapping of bacterial cytoskeletal mutants reveals physiological robustness to cell size. Current Biology 27, 3419-3429. e3414 (2017).
- A. Colavin, H. Shi, K. C. Huang, RodZ modulates geometric localization of the bacterial actin MreB to regulate cell shape. Nature communications 9, 1280 (2018).
- L. K. Harris, J. A. Theriot, Relative rates of surface and volume synthesis set bacterial cell size. Cell 165, 1479-1492 (2016).
- C. Brownlee, R. Heald, Importin α partitioning to the plasma membrane regulates intracellular scaling. Cell 176, 805-815. e808 (2019).
Follow the Topic
-
Nature Communications
An open access, multidisciplinary journal dedicated to publishing high-quality research in all areas of the biological, health, physical, chemical and Earth sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Women's Health
Publishing Model: Hybrid
Deadline: Ongoing
Tumor Microenvironment Crosstalk and Therapeutic Implications
Publishing Model: Hybrid
Deadline: Nov 02, 2026
Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in