The resilience of marine organisms to overshoot global climate
Published in Earth & Environment
Anthropogenic climate change threats marine organisms by altering their environment (IPCC 2021, IPCC 2022). Indeed, the combined effects of ocean warming, deoxygenation (decrease in oxygen content) and acidification (increase in pH) -the so-called "deadly trio" (Bjima et al., 2013)- are already impacting dramatically the organisms of the oceans, and will continue to do so with ongoing global warming.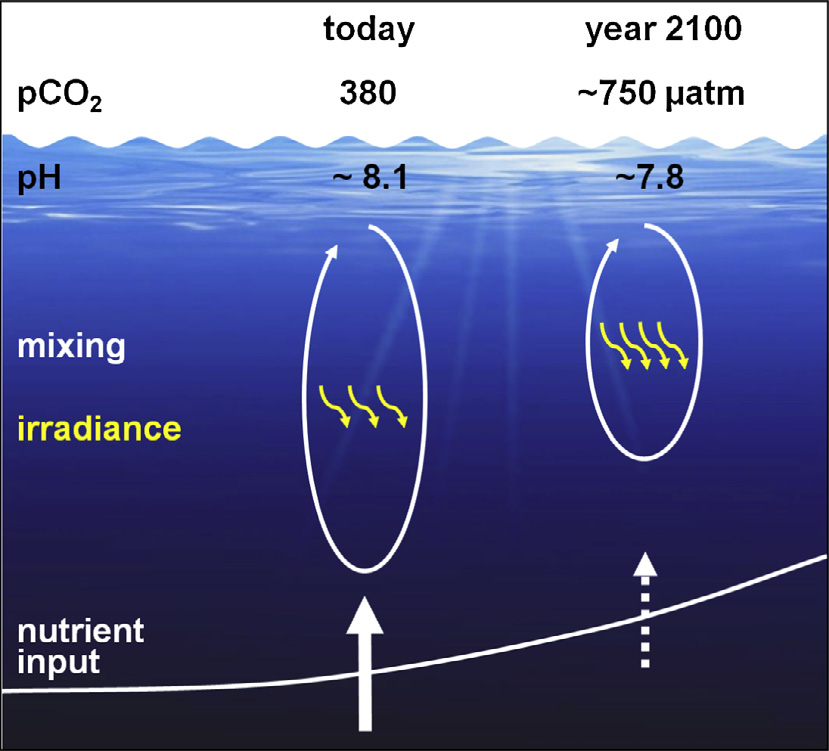
From Bjima et al. (2013). Figure summarizing main changes in the ocean during this century; intensification of the ocean acidification, ocean warming that will affect vertical mixing and, thus, the nutrient inputs from deeper layers.
To mitigate these and other undesired effects, almost 200 countries reaffirmed at Glasgow COP26 (2021) the agreement signed at Paris COP21 (2015) to limit global warming to 1.5ºC, and no more than 2º C. Nonetheless, recent trends in global greenhouse gases (GHG) emissions suggest this limit will be exceeded in the following decades (Mathews and Wynes 2022). Stabilising the climate would require ongoing net-zero GHG emissions, yet returning to lower temperature levels if those limits are crossed would require the massive deployment of technologies for direct removal of the excess atmospheric carbon dioxide (CDR; Shepherd 2009). However, exceeding these temperature thresholds will come with a cost.
Several studies have investigated the response of Earth systems to exceeding this limit and to, eventually, get back on track afterwards, that is, an overshoot. In the oceans, it has been suggested that the deadly trio may recover in the upper oceans if carbon dioxide concentrations were to decrease (Frölicher and Joos 2010); yet changes on them persist far into the future in the ocean interior (Bertini and Tjiputra 2022). However, the persistence of their combined effects to marine organisms after an eventual overshoot is yet to be investigated.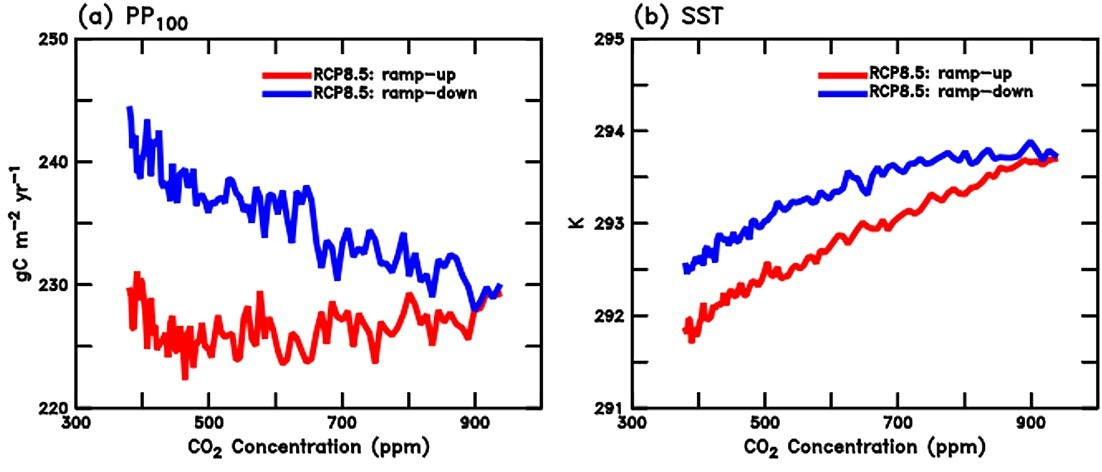 From John et al. (2015). Figure showing global ocean mean 100 m depth primary production and sea surface temperature during a concentration-driven ramp-up/ramp-down experiment. Both parameters present slight hysteresis at the end of the experiment with respect to the initial conditions.
From John et al. (2015). Figure showing global ocean mean 100 m depth primary production and sea surface temperature during a concentration-driven ramp-up/ramp-down experiment. Both parameters present slight hysteresis at the end of the experiment with respect to the initial conditions.
In this framework, my co-authors and I have quantitatively accounted for the combined effects of ocean warming and oxygen loss in the ability of the world oceans to support viable ecosystems by using the metabolic index (φ, Deutsch et al., 2015). This index brings together global warming perturbations with their effects on the ocean distribution of life by accounting for the ratio of oxygen supply to resting demand; both of which depending on ambient partial pressure of oxygen, temperature, and species-specific physiological terms. In particular, we have estimated this metabolic index for 72 different species throughout two experiments that simulate global temperature overshoots on seven Earth System models. Specifically, we have used an idealised experiment (1pctCO2) that simulates a rapid increase of atmospheric carbon dioxide concentrations from pre-industrial levels to four times initial values, and then mirrored decline, and (2) a scenario (SSP5-3.4) in which GHG emissions continue up to 2040 at which point aggressive mitigation is assumed.
We observed that φ is positively/negatively correlated with oxygen partial pressure/temperature at the sea surface throughout the full experiments, though its hysteresis (i.e., the difference of the final state with respect to initial conditions) is higher than that of its drivers in isolation. As we get deeper into the dark ocean, the pathway of φ differs from those of oxygen partial pressure and temperature showing a legacy of concomitant changes in its drivers on ocean habitability. In addition, the hysteresis of the three parameters increase in agreement with recent literature.
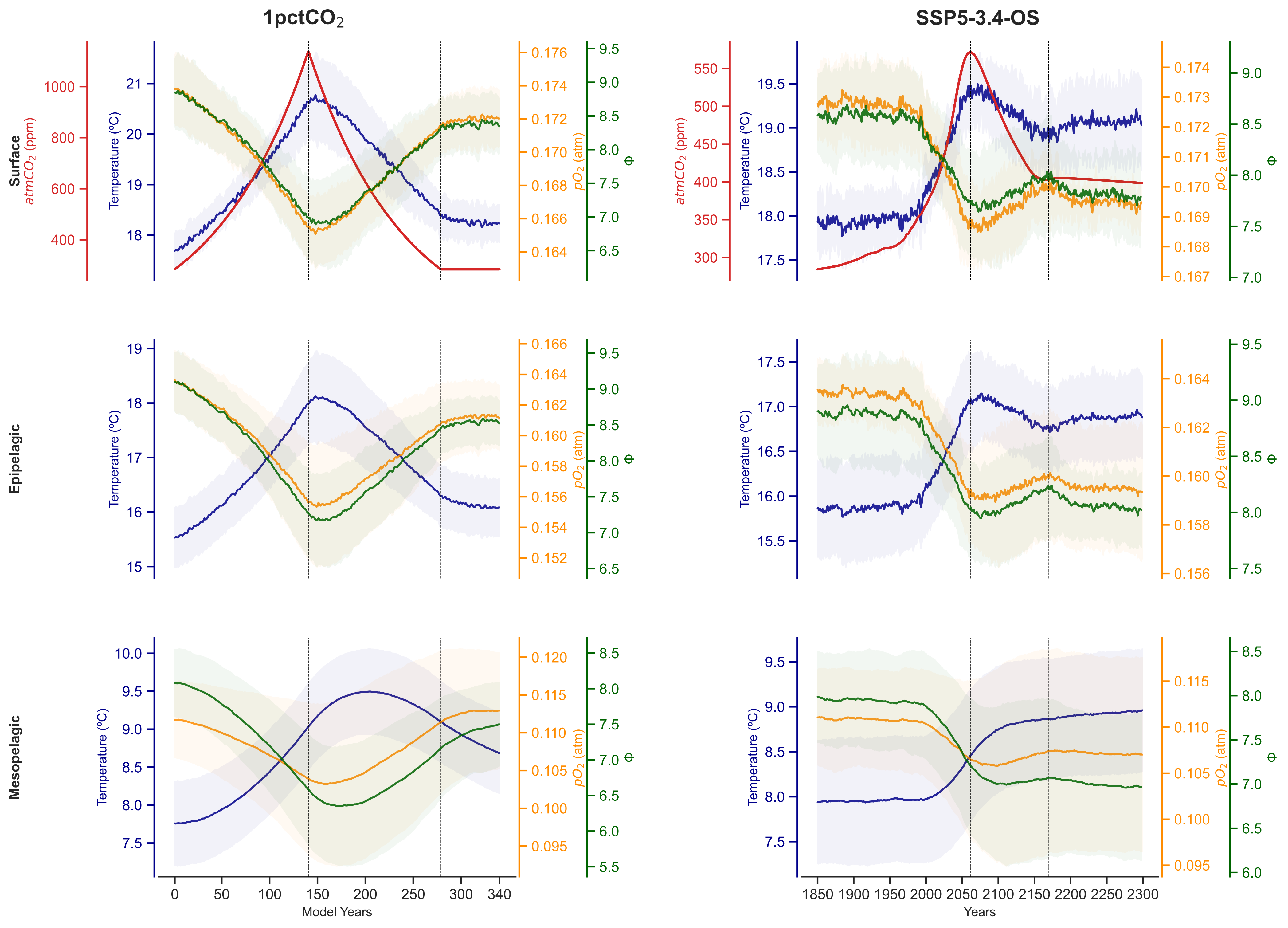
At the sea surface, the metabolic index (in green) follows the evolution of partial pressure of oxygen (in orange) during both experiments, which is inversely correlated with changes in sea surface temperature (in blue). In deeper waters, the metabolic index moves away from the behaviour of partial pressure of oxygen and temperature.
When analyzing the timescale of those changes after global temperature recovers from the overshoot, we observed that the combined impacts of ocean warming and deoxygenation, as measured by φ, will last longer than the changes on them in isolation.
The metabolic index also allows us to differentiate between ocean ecosystems that can hold viable habitats from those that are unsuitable for a particular organism to thrive. In particular, for an organism to carry out aerobic metabolic activities, oxygen supply must meet at minimum the resting metabolic demand (that is, φ=1); though additional energy (that is, φ>>1) needs to be undertaken to accomplish key ecological activities like feeding, growth and reproduction. These "critical" values has been observed to vary within 2 to 7 in the ocean environments. In our study, we have followed the evolution of the global ocean volume at which φ is below the resting rate (non-viable zone), between the resting and those critical values (critical zone), and above those critical values (viable zone). 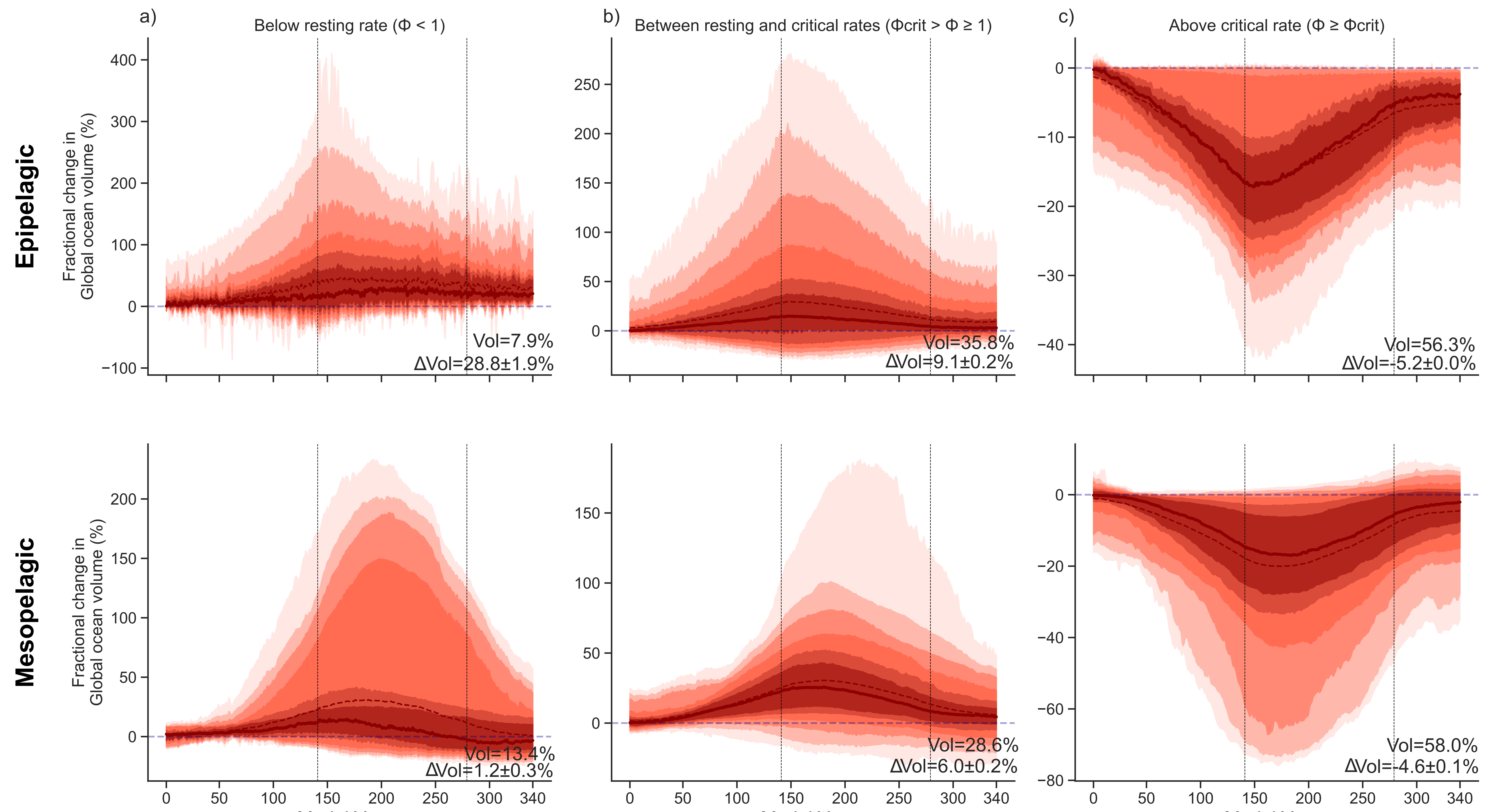
During the 1pctCO2 experiment, the volume of waters that cannot hold viable habitats increase after the experiment with respect to the initial conditions. In contrast, the volume of waters in which conditions are suitable for viable habitats decrease after the overshoot.
Consistently across the seven models, and the two overshoot experiments, the volume of waters that can hold viable habitats decreases after global temperature recovers from the overshoot with respect to the initial state. This decrease occurs at the expense of the volume of waters that are unable to sustain viable habitats that increases.
Our study suggests that the alterations that global warming induces to the world ocean will decrease its capacity for providing viable habitats for marine organisms. These alterations are likely to require centuries to recover even if the global temperature returns back to initial levels. As our analysis considers relatively rapid increase and decrease of global temperature, it is expected that a "real" overshoot and implementation of negative emissions following CDR deployment would take much longer. Overall, this study encourages us to the prompt implementation of mitigation actions as delaying climate stabilisation is highly likely to impact irreversibly marine ecosystems.
References:
Bertini, L., and Tjiputra, J. Biogeochemical timescales of climate change onset and recovery in the North Atlantic interior under rapid atmospheric CO2 forcing. JGR Oceans 127 e2021JC017929
Bijma et al. Climate change and the oceans - What does the future hold? Marine Pollution Bulletin 74, 495-505 (2013)
IPCC, 2021: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assesment Report of the Intergovernmental Panel on Climate Change [Masson-Delmotte, V., et al. (eds). Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA.
Deutsch et al. Climate change tightens a metabolic constraint on marine habitats. Science 348, 1132-1135 (2015).
IPCC, 2022: Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [H.-O. Pörtner, et al. (eds.)]. Cambridge University Press. Cambridge University Press, Cambridge, UK and New York, NY, USA, 3056 pp.
Frölicher, T.L., Joos, F. Reversible and irreversible impacts of greenhouse gas emissions in multi-century projections with the NCAR global coupled carbon cycle-climate model. Climate Dynamics 35, 1439–1459 (2010).
John, J.G., Stock, C.A., and Dunne, J.P. A more productive, but different, ocean after mitigation. Geophysical Research Letters 42, 9836-9845 (2015).
I'm a postdoc researcher at CNRM-MétéoFrance investigating how global warming induced changes in the ocean may affect marine ecosystems.
Follow the Topic
-
Communications Earth & Environment
An open access journal from Nature Portfolio that publishes high-quality research, reviews and commentary in the Earth, environmental and planetary sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Hazards in Mountain Regions
Publishing Model: Open Access
Deadline: Nov 02, 2026
Ecosystems under marine heatwaves
Publishing Model: Hybrid
Deadline: Oct 22, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in