The story behind the Nature Microbiology article “Immunogenicity of poxvirus A16/G9 entry-fusion subcomplex and its restriction by A56/K2 protein informs vaccine design”
Published in Biomedical Research
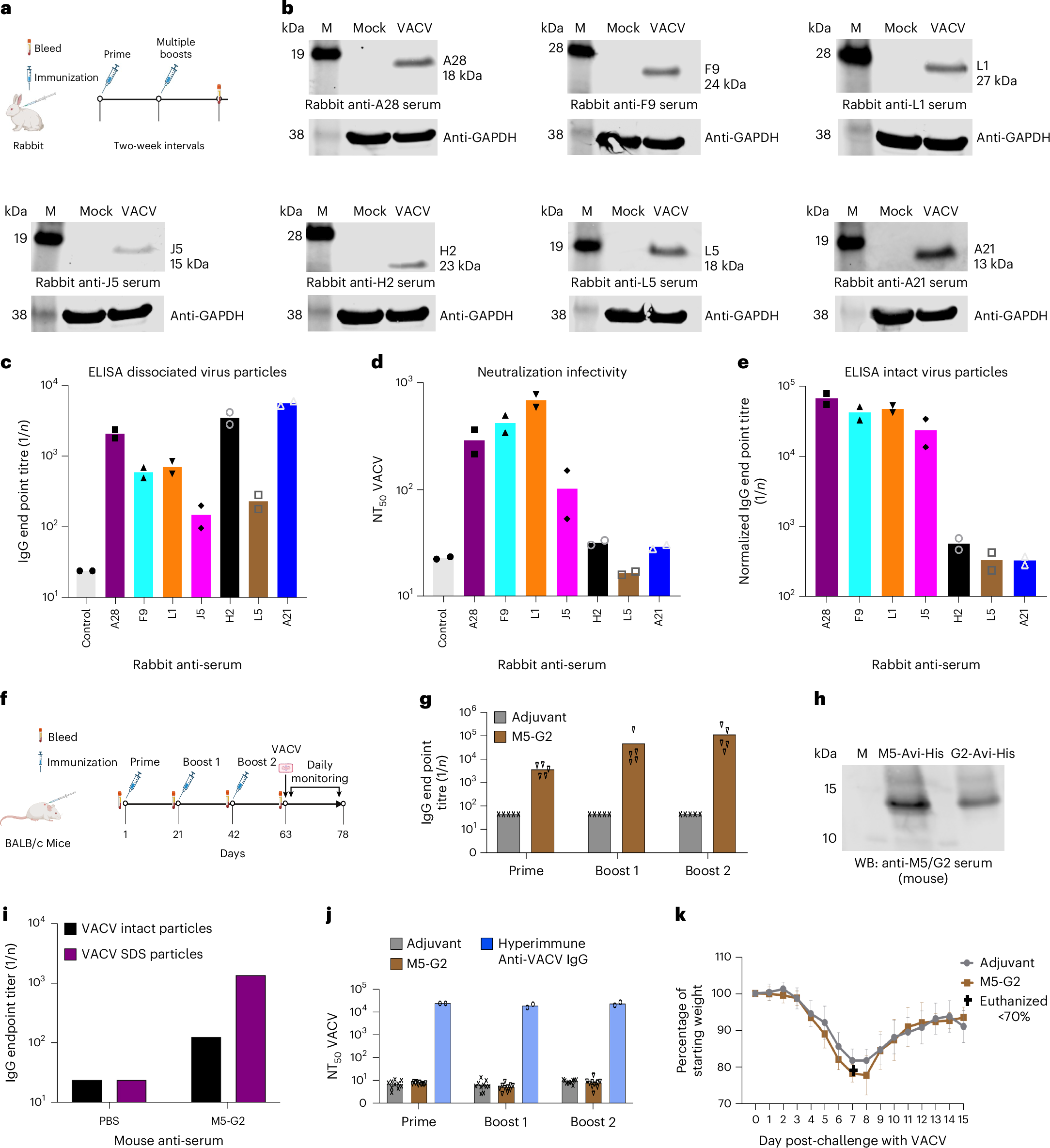
The story begins more than 20 years ago. In 2004, Christiana Fogg – a University of Maryland graduate student doing her thesis research in my NIH lab, Shlomo Lustig – a visiting scholar from the Israel Institute for Biological Research, and Gary Cohen – a researcher at the University of Pennsylvania joined forces to investigate the requirements for protection against orthopoxvirus infection in an animal model. At the time it was known that there are two forms of infectious vaccinia virus (VACV) – the mature virion (MV) and the extracellular virion (EV). The MV is present intracellularly and is also enclosed by the outer membrane of the EV. We established that antibodies to proteins of both the MV and outer EV membrane are required to completely protect mice against weight loss following a lethal VACV challenge 1, 2.
In that same year (2004) Tatiana Senkevich – a staff scientist in my lab identified the first poxvirus entry-fusion protein 3. The discovery of poxviral entry proteins had been delayed for several reasons. VACV, the model orthopoxvirus, can enter mammalian, avian, amphibian and some insect cells suggesting there is no unique cellular receptor. Furthermore, over 40 proteins were thought to be membrane-associated based on predicted amino acid sequences and experimental data. To reduce the number of candidate entry proteins, we prioritized those that are highly conserved in all poxviruses for construction of conditional lethal mutants. Tatiana, together with postdoctoral fellows and students eventually identified 11 proteins in the MV membrane that are each required for entry and form the entry-fusion complex (EFC) 4-11. In 2008, Tim Wagenaar, another University of Maryland graduate student doing thesis research in my lab, discovered that a complex comprised of the A56 and K2 proteins interacted specifically with the A16/G9 EFC subcomplex 12. A56 and K2 are expressed late in infection and inserted into the plasma membrane. When the genes encoding A56/K2 are deleted, progeny virions on the surface of the parental cell fuse back depositing the EFC in the plasma membrane leading to formation of giant syncytia 13. Thus, A56/K2 prevents superinfection and acts as a virus and cell fusion-suppressor.
The 2022 global outbreak of mpox, caused by monkeypox virus (MPXV), stimulated us to systematically analyze the immunogenicity of the EFC proteins to aid in the development of improved vaccines. Although each EFC protein is a potential antibody target, only two of the eleven had been shown to induce protective immunity. In 2024, Huibin Yu – a postdoctoral fellow took the lead on this project. The work was jump-started with seven purified EFC proteins that Gary Cohen had in his freezer and Huibin synthesized and purified the additional ones. Of the ten proteins analyzed (the small 35-amino acid O3 component consisting mostly of a transmembrane domain was not analyzed), six induced neutralizing antibodies. Wolfgang Resch, an expert in bioinformatics in the NIH Center for Information Technology (and a former postdoctoral fellow in my lab), modeled the EFC initially using AlphaFold2 and then upgrading the model with AlphaFold3. The ectodomains of non-neutralizing targets were located near the base with the A16/G9 subcomplex forming the apex. Focusing on A16/G9, we demonstrated that it induced potent orthopoxvirus cross-neutralizing protective antibodies. An unexpected finding was that little or no antibodies to A16/G9 were detected in serum from vaccinees that received the licensed smallpox/mpox vaccine or animals that were infected with VACV, MPXV or cowpox virus. Our previous studies had shown that A16 and G9 are more firmly embedded in the viral membrane than other EFC proteins as they are not released from virus particles by non-ionic detergents, offering a partial explanation for their poor immunogenicity. We submitted the manuscript to Nature Microbiology with the thought of addressing in future studies the apparent paradox of how A16/G9 can be targets of neutralizing antibodies but not immunogenic in live virus vaccines. However, following the mostly favorable comments of the Reviewers and Editor, we moved up our plan and over the ensuing six months of revision determined that the A56/K2 complex has an additional role in suppressing the immunogenicity of A16/G9. VACV with a deletion of the A56/K2 genes made antibodies to A16/G9 after a single immunization, which were boosted after a second immunization. Our current model is that B-cells in lymph nodes can access A16/G9 when inserted into the plasma membrane (which occurs when A56/K2 is not expressed) or when secreted but not when A16/G9 is sequestered in the virus particle. In contrast to immobilized membrane bound B-cell receptors, flexible circulating antibodies have a window of opportunity to neutralize MVs when the outer EV membrane is disrupted to allow fusion with cellular membranes.
Our findings could lead to improved vaccines in several possible ways. One is by expressing A16/G9 along with other MV and EV antigens in recombinant mRNA or protein nanoparticle vaccines. Another is by modifying live attenuated vaccines to express secreted forms of A16/G9 or by deleting A56 and/or K2.
References
- Fogg, C. et al. Protective immunity to vaccinia virus induced by vaccination with multiple recombinant outer membrane proteins of intracellular and extracellular virions. J. Virol. 78, 10230–10237 (2004).
- Lustig, S., Fogg, C., Whitbeck, J.C. & Moss, B. Synergistic neutralizing activities of antibodies to outer membrane proteins of the two infectious forms of vaccinia virus in the presence of complement. Virology 328, 30–35 (2004).
- Senkevich, T.G., Ward, B.M. & Moss, B. Vaccinia virus entry into cells is dependent on a virion surface protein encoded by the A28L gene. J. Virol. 78, 2357–2366 (2004).
- Senkevich, T.G. & Moss, B. Vaccinia virus H2 protein is an essential component of a complex involved in virus entry and cell-cell fusion. J. Virol. 79, 4744–4754 (2005).
- Senkevich, T.G., Ojeda, S., Townsley, A., Nelson, G.E. & Moss, B. Poxvirus multiprotein entry-fusion complex. Proc. Natl. Acad. Sci. USA 102, 18572–18577 (2005).
- Townsley, A., Senkevich, T.G. & Moss, B. Vaccinia virus A21 virion membrane protein is required for cell entry and fusion. J. Virol. 79, 9458–9469 (2005).
- Ojeda, S., Senkevich, T.G. & Moss, B. Entry of vaccinia virus and cell-cell fusion require a highly conserved cysteine-rich membrane protein encoded by the A16L gene. J. Virol. 80, 51–61 (2006).
- Brown, E., Senkevich, T.G. & Moss, B. Vaccinia virus F9 virion membrane protein is required for entry but not virus assembly, in contrast to the related l1 protein. J. Virol. 80, 9455–9464 (2006).
- Izmailyan, R.A., Huang, C.Y., Mohammad, S., Isaacs, S.N. & Chang, W. The envelope G3L protein is essential for entry of vaccinia virus into host cells. J. Virol. 80, 8402–8410 (2006).
- Ojeda, S., Domi, A. & Moss, B. Vaccinia virus G9 protein is an essential component of the poxvirus entry-fusion complex. J . Virol. 80, 9822–9830 (2006).
- Bisht, H., Weisberg, A.S. & Moss, B. Vaccinia virus L1 protein is required for cell entry and membrane fusion. J. Virol. 82, 8687–8694 (2008).
- Wagenaar, T.R., Ojeda, S. & Moss, B. Vaccinia virus A56/K2 fusion regulatory protein interacts with the A16 and G9 subunits of the entry fusion complex. J. Virol. 82, 5153–5160 (2008).
- Wagenaar, T.R. & Moss, B. Expression of the A56 and K2 proteins is sufficient to inhibit vaccinia virus entry and cell fusion. J. Virol. 83, 1546–1554 (2009).
This research was supported by the Intramural Research Program of the National Institutes of Health (NIH). The contributions of the NIH author are considered Works of the United States Government. The findings and conclusions presented in this paper are those of the author and do not necessarily reflect the views of the NIH or the U.S. Department of Health and Human Services.
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in