WIPI1 enhances Autophagy and Longevity
Published in Cell & Molecular Biology
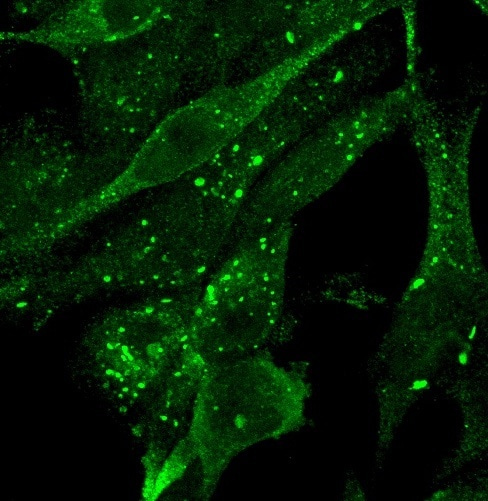
Many years ago, we discovered the human WIPI protein family, whose four members, WIPI1 to WIPI41, exert important PI3P effector functions in the process of autophagy2. In fact, by examining WIPI1 back then, we were able for the first time to assign a role for human WIPI proteins, then novel 7-bladed beta-propeller proteins, in starvation-induced autophagy and cancer1. In this context, we observed early on that WIPI1 accumulates during autophagy induction in the perinuclear region1 where it appeared to be associated with the ER (Video 1), however, the underlying mechanism for this feature remained unclear. Later, in a comprehensive and comparative analysis of the function of all four human WIPI proteins, we could show that WIPI1 interacts with WIPI2 through protein-protein interaction3. WIPI2 performs a scaffold function on the nascent autophagosome (also referred to as phagophore), whereby WIPI2 binds to PI3P of the autophagosomal membrane and via specific protein-protein interaction recruits the ATG16L1 protein complex, which is important for autophagosome formation3-5. We assumed that WIPI1 supports the WIPI2 function here, but the specific function of WIPI1 still remained unclear.
Video 1. Typical perinuclear GFP-WIPI1 localization during starvation-induced autophagy initiation. The video focuses on and magnifies the perinuclear (nucleus in blue) colocalization of GFP-WIPI1 (in green) and the ER (in red) in human U2OS cells (ATCC). U2OS cells stably expressing GFP-WIP11 cells were treated with nutrient-free medium (EBSS) for 3 h, fixed with 3.7 % paraformaldehyde, stained using anti-ERp61 (1:25, BD Biosciences) and anti-mouse IgG Alexa 546 (1:250, Life Technologies) antibodies as well as TO-PRO-3 (1:1000, Life Technologies). Confocal laser-scanning microscopy was performed with a Zeiss LSM 510 microscope using a 63x/1.4 DIC Plan-apochromat objective, and Z-stack images acquired with 19 optical sections of 0.2 µm and a resolution of 512 x 512 pixels per individual image. Using Volocity 3.1 software (Improvision), Z-stack images were used for 3D-reconstruction and fly-through movie presentations. This experiment was carried out as part of Simon G. Pfisterer's doctoral thesis in the Proikas Cezanne Laboratory at the Eberhard Karls University in Tübingen.
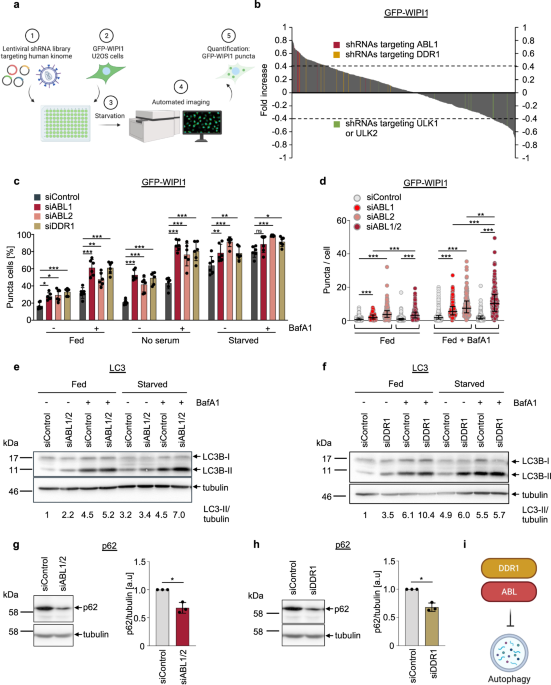
To question the specific function of WIPI1, we considered whether identifying regulatory pathways of WIPI1 might help us here. At that time, we had established an automated method for quantitative, fluorescence-based WIPI1 image acquisition and analysis6. Based on this, we used human U2OS cells expressing the fluorescent GFP-tagged WIPI1 protein and performed an unbiased screen to identify protein kinases that could regulate WIPI1. We have identified ABL1 (and ABL2) and DDR1 protein kinases and, in combination with further unbiased studies using quantitative phosphoproteomics and quantitative gene expression analysis, we were able to decode the following scenario in the context of WIPI1 regulation.
WIPI1 acts as an enhancer of autophagy and is kept at a low basal gene expression level by the ABL1/2-ERK2-MYC signalling axis (Figure 1). Thereby, the transcription factor MYC binds to the WIPI1 promotor and represses WIPI1 gene expression. Counteracting ABL1/2-ERK2-MYC signalling and subsequently increasing WIPI1 levels enhances autophagy. In the context of its function as an enhancer of autophagy, we were able to show that the typical WIPI1 accumulation in connection with the ER (Video 1) represents the formation of several phagophores. According to this, WIPI1 seems to exercise its enhancer effect by stimulating the formation of more phagophores and thus ultimately more autophagosomes. Our future task will now be to decipher how this stimulation is mediated on a molecular level.

Figure 1. Word cloud for regulation of WIPI1 in autophagy with implications for life span control in C. elegans. Keywords (WIPI1, autophagy, lifespan, ABL1, ERK2, MYC, gene expression, TNTs) were derived from our study and used to create a word cloud with WordArt.com, using the recycling symbol to visually reference the process of autophagy.
During our characterization of WIPI1 as an autophagy enhancer, we provide evidence, that autophagy material can be sent from one cell to another cell via tunneling nanotubes7 (TNTs), and that the amount of WIPI1 can also influence this transport here. This finding is currently the least understood molecularly since the importance of TNTs in general in vivo is still unclear. However, we found it interesting to investigate this finding in the context of WIPI1 and were able to show that the transport of autophagy material is increased into cells that have low autophagy activity. We hypothesize that this could be a compensatory way in which cells support each other to adequately respond to stressful situations through autophagy.
When we carried out the screening to identify WIPI1 regulatory protein kinases, we also established the model organism C. elegans in the laboratory8 to further validate our results in a physiological context, and to conduct investigations into the connection between autophagy and longevity. For us, this was particularly interesting after our new finding on the regulation of WIPI1 by ABL1/2-ERK2-MYC signalling for the following reasons. First, it was shown early in the model organism C. elegans that regulation of lifespan depends on autophagy9,10. Therefore, we hypothesized that the ABL1/2-ERK2-MYC signaling axis, which inhibits WIPI1 and autophagy in human cells, should prolong the lifespan of C. elegans when counteracted. Indeed, our respective studies showed that the absence of ABL prolongs the lifespan of C. elegans and that this depends on the WIPI1 homolog in C. elegans, which was expressed at a higher level without ABL, while autophagic flux was also increased.
This result, albeit on a speculative basis for the time being, can now be viewed in the following context of human longevity. Centenarians appear to have elevated levels of WIPI1 expression11. Therefore, there might be an advantage if the ABL-ERK-MYC signalling axis is less effective with age to increase the expression of WIPI1 and thereby increase its function as an enhancer of autophagy. So, would it be a good idea to increase expression of WIPI1 with age, as seems to be the case in centenarians11, to gently increase autophagy? Well, we don't know (yet).
In summary, our study raises three important directions for future research:
- How does WIPI1 fulfill its enhancer function in forming multiple phagophores at PI3P-enriched ER regions? Does this occur through WIPI1 oligomerization?
- How is the ABL1/2-ERK2-MYC signalling axis that controls WIPI1 gene expression regulated? In our study, we indicate that the receptor tyrosin-specific protein kinase DDR1 is located upstream of ABL1/2. How is the signalling between DDR1 and ABL1/2 executed?
- Which mechanism regulates the transport of autophagy material between cells by TNTs and which specific role does WIPI1 play here? Does TNT transport of autophagy material exist in vivo and what are the physiological consequences?
References
1 Proikas-Cezanne, T. et al. WIPI-1alpha (WIPI49), a member of the novel 7-bladed WIPI protein family, is aberrantly expressed in human cancer and is linked to starvation-induced autophagy. Oncogene 23, 9314-9325 (2004). https://doi.org:10.1038/sj.onc.1208331
2 Dikic, I. & Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat Rev Mol Cell Biol 19, 349-364 (2018). https://doi.org:10.1038/s41580-018-0003-4
3 Bakula, D. et al. WIPI3 and WIPI4 beta-propellers are scaffolds for LKB1-AMPK-TSC signalling circuits in the control of autophagy. Nat Commun 8, 15637 (2017). https://doi.org:10.1038/ncomms15637
4 Polson, H. E. et al. Mammalian Atg18 (WIPI2) localizes to omegasome-anchored phagophores and positively regulates LC3 lipidation. Autophagy 6, 506-522 (2010). https://doi.org:10.4161/auto.6.4.11863
5 Dooley, H. C. et al. WIPI2 links LC3 conjugation with PI3P, autophagosome formation, and pathogen clearance by recruiting Atg12-5-16L1. Mol Cell 55, 238-252 (2014). https://doi.org:10.1016/j.molcel.2014.05.021
6 Pfisterer, S. G., Mauthe, M., Codogno, P. & Proikas-Cezanne, T. Ca2+/calmodulin-dependent kinase (CaMK) signaling via CaMKI and AMP-activated protein kinase contributes to the regulation of WIPI-1 at the onset of autophagy. Mol Pharmacol 80, 1066-1075 (2011). https://doi.org:10.1124/mol.111.071761
7 Zurzolo, C. Tunneling nanotubes: Reshaping connectivity. Curr Opin Cell Biol 71, 139-147 (2021). https://doi.org:10.1016/j.ceb.2021.03.003
8 Takacs, Z. et al. ATG-18 and EPG-6 are Both Required for Autophagy but Differentially Contribute to Lifespan Control in Caenorhabditis elegans. Cells 8 (2019). https://doi.org:10.3390/cells8030236
9 Melendez, A. et al. Autophagy genes are essential for dauer development and life-span extension in C. elegans. Science 301, 1387-1391 (2003). https://doi.org:10.1126/science.1087782
10 Toth, M. L. et al. Longevity pathways converge on autophagy genes to regulate life span in Caenorhabditis elegans. Autophagy 4, 330-338 (2008). https://doi.org:10.4161/auto.5618
11 Xiao, F. H. et al. Transcriptome evidence reveals enhanced autophagy-lysosomal function in centenarians. Genome Res 28, 1601-1610 (2018). https://doi.org:10.1101/gr.220780.117
Abbreviations
ABL1: ABL proto-oncogene 1; non-receptor tyrosine-specific protein kinase
ABL2: ABL proto-oncogene 2; non-receptor tyrosine-specific protein kinase
DDR1: Discoidin Domain Receptor Tyrosine Kinase 1; receptor tyrosine-specific protein kinase
ERK2: Extracellular signal-regulated kinase 2; MAP kinase
MYC: MYC proto-oncogene; transcription factor
ATG16L1: Autophagy related 16L1; autophagy gene
PI3P: Phosphatidylinositol 3-phosphate; phospholipid
TNT: Tunneling nanotubule; cell-to-cell connection
WIPI: WD-repeat protein interacting with phosphoinositides; 7-bladed beta-propeller protein
Follow the Topic
-
Communications Biology

An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.
Related Collections
With Collections, you can get published faster and increase your visibility.
Forces in Cell Biology
Publishing Model: Open Access
Deadline: Apr 30, 2026
From RNA Detection to Molecular Mechanisms
Publishing Model: Open Access
Deadline: May 05, 2026



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in