Elevated ATGL in colon cancer cells and cancer stem cells promotes metabolic and tumorigenic reprogramming reinforced by obesity
Published in Cancer
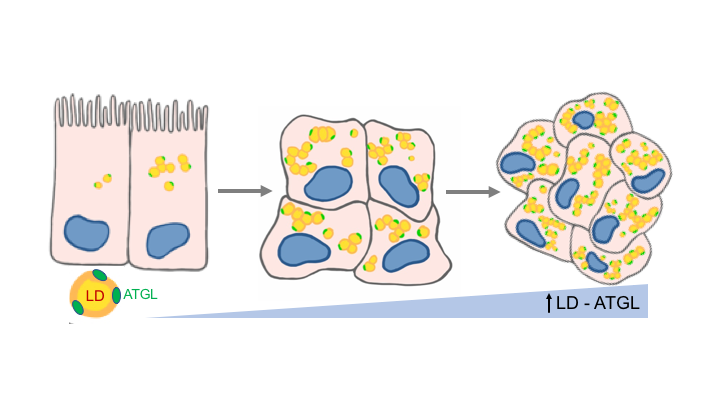

Obesity, increasingly prevalent in both industrialized and developing countries, is estimated to affect 1.6 billion people by 2030 (1). This global health crisis is associated with various diseases such as diabetes, cardiovascular disorders, inflammatory disorders, and cancer (2). Obesity increases the risk and progression of colon cancer, the second leading cause of cancer-related death worldwide (3, 4). Colon cancer patients with obesity have prevalent tumor recurrence and resistance to chemotherapy (5). Therefore, there is an excessive, unmet demand to understand how obesity facilitates colon cancer development and recurrence and to develop more effective treatment options.
Human cancers, including colon cancer, are characterized by systemic metabolic changes needed to provide fuel for tumor cells’ excessive growth and migration to distant organs (6, 7). These metabolic changes involve energy sources from organelles such as lipid droplets (LDs) which are warehouses for intracellular lipids (8, 9). Our group, and others, have demonstrated that LDs, substantially elevated in colonic tumors, are important for tumor growth, and emerging findings suggest that they may be critical for cancer stem cell behavior (10-13). Further, obesity mediators and metabolites stimulate LDs accumulation in colon cancer cells (14, 15). As LD utilization is mediated exclusively by adipose triglyceride lipase (ATGL), also known as PNPLA2 (16), we hypothesized that obesity increases ATGL-mediated LDs utilization, thus facilitating colon cancer progression through metabolic and oncogenic changes in colon cancer cells and colon cancer stem cells.
We found that increased ATGL levels in local colon cancer patients’ tumors, compared to controls, were further augmented in obese individuals. Elevated ATGL levels in human colon cancer cells were also augmented by an obesity mediator. Further, inhibiting ATGL blocked LDs utilization in colon cancer cells, leading to the attenuation of cancer cell growth and migration. Ultimately, inhibition of ATGL effectively attenuated growth of colon cancer stem cells. This is important as resistance to therapy and cancer recurrence in colon cancer patients, both of which are prevalent in obese individuals, are mainly due to the heterogenic nature and regenerative potential of colon cancer stem cells (5, 17). Therefore, elevated ATGL-mediated LD utilization in colon cancer cells may be critical in driving cancer progression, especially in conjunction with obesity. In addition, this may be a common characteristic among heterogenic colon cancer stem cells populations, thus making ATGL a promising therapeutic target.
Next, we found that ATGL inhibition targets the expression of about five thousand genes in colon cancer cells and colon cancer stem cells that are drivers of cancer, gastrointestinal and metabolic diseases, as well as biological processes related to metabolism and growth. We further identified several ATGL-dependent genes in colon cancer cells and colon cancer stem cells whose roles in cancer progression are not well understood. For example, ATG2B regulates autophagosome formation and LDs morphology (18), while PCK2 is involved in the citric acid cycle (19). Further, SPTLC2 regulates sphingolipid synthesis (20), while PGAM1 is involved in glycolysis (21). These data revealed that ATGL mediated systemic changes in colon cancer cells and cancer stem cells are linked to oncogenic process and a cross-talk between different sources of energy in colon cancer cells and colon cancer stem cells.
Colon cancer incidence, especially in young adults, has been steadily increasing with the rising rates of obesity (3, 4), which points to an urgent, unmet demand to understand these processes. Here, we demonstrated one of the mechanisms through which obesity may reinforce colon cancer progression involved ATGL-mediated utilization of LDs, which are elevated in colon cancer cells and colon cancer stem cells. This provides a novel understanding of the interplay between oncogenic and metabolic processes, which will allow us to establish a platform for new diagnostic and therapeutic approaches for colon cancer patients especially in conjunction with obesity.
Reference
- Popkin BM, Adair LS, Ng SW. Global nutrition transition and the pandemic of obesity in developing countries. Nutrition reviews. 2012;70(1):3-21.
- Fruh SM. Obesity: Risk factors, complications, and strategies for sustainable long-term weight management. J Am Assoc Nurse Pract. 2017;29(S1):S3-S14.
- Rawla P, Sunkara T, Barsouk A. Epidemiology of colorectal cancer: incidence, mortality, survival, and risk factors. Prz Gastroenterol. 2019;14(2):89-103.
- Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: a cancer journal for clinicians. 2018;68(6):394-424.
- Ayob AZ, Ramasamy TS. Cancer stem cells as key drivers of tumour progression. J Biomed Sci. 2018;25(1):20.
- Zhu X, Chen HH, Gao CY, Zhang XX, Jiang JX, Zhang Y, et al. Energy metabolism in cancer stem cells. World J Stem Cells. 2020;12(6):448-61.
- Sanchez-Martinez R, Cruz-Gil S, Gomez de Cedron M, Alvarez-Fernandez M, Vargas T, Molina S, et al. A link between lipid metabolism and epithelial-mesenchymal transition provides a target for colon cancer therapy. Oncotarget. 2015;6(36):38719-36.
- Welte MA, Gould AP. Lipid droplet functions beyond energy storage. Biochim Biophys Acta Mol Cell Biol Lipids. 2017;1862(10 Pt B):1260-72.
- Petan T. Lipid Droplets in Cancer. Rev Physiol Biochem Pharmacol. 2020.
- Accioly MT, Pacheco P, Maya-Monteiro CM, Carrossini N, Robbs BK, Oliveira SS, et al. Lipid bodies are reservoirs of cyclooxygenase-2 and sites of prostaglandin-E2 synthesis in colon cancer cells. Cancer Res. 2008;68(6):1732-40.
- Straub BK, Herpel E, Singer S, Zimbelmann R, Breuhahn K, Macher-Goeppinger S, et al. Lipid droplet-associated PAT-proteins show frequent and differential expression in neoplastic steatogenesis. Mod Pathol. 2010;23(3):480-92.
- Qi W, Fitchev PS, Cornwell ML, Greenberg J, Cabe M, Weber CR, et al. FOXO3 growth inhibition of colonic cells is dependent on intraepithelial lipid droplet density. J Biol Chem. 2013;288(23):16274-81.
- Alvina FB, Gouw AM, Le A. Cancer Stem Cell Metabolism. Advances in experimental medicine and biology. 2021;1311:161-72.
- Fazolini NP, Cruz AL, Werneck MB, Viola JP, Maya-Monteiro CM, Bozza PT. Leptin activation of mTOR pathway in intestinal epithelial cell triggers lipid droplet formation, cytokine production and increased cell proliferation. Cell Cycle. 2015;14(16):2667-76.
- Heller S, Cable C, Penrose H, Makboul R, Biswas D, Cabe M, et al. Intestinal inflammation requires FOXO3 and Prostaglandin E2 dependent lipogenesis and elevated lipid droplets. Am J Physiol Gastrointest Liver Physiol. 2016:ajpgi 00407 2015.
- Zimmermann R, Strauss JG, Haemmerle G, Schoiswohl G, Birner-Gruenberger R, Riederer M, et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science. 2004;306(5700):1383-6.
- DeClercq V, McMurray DN, Chapkin RS. Obesity promotes colonic stem cell expansion during cancer initiation. Cancer letters. 2015;369(2):336-43.
- Klionsky DJ. Autophagy: from phenomenology to molecular understanding in less than a decade. Nat Rev Mol Cell Biol. 2007;8(11):931-7.
- Shin HD, Park BL, Kim LH, Cheong HS, Kim JH, Cho YM, et al. Association of a polymorphism in the gene encoding phosphoenolpyruvate carboxykinase 1 with high-density lipoprotein and triglyceride levels. Diabetologia. 2005;48(10):2025-32.
- Hojjati MR, Li Z, Jiang XC. Serine palmitoyl-CoA transferase (SPT) deficiency and sphingolipid levels in mice. Biochim Biophys Acta. 2005;1737(1):44-51.
- Hitosugi T, Zhou L, Elf S, Fan J, Kang HB, Seo JH, et al. Phosphoglycerate mutase 1 coordinates glycolysis and biosynthesis to promote tumor growth. Cancer cell. 2012;22(5):585-600.
Follow the Topic
-
Oncogenesis
A peer-reviewed open access online journal that publishes articles exploring mechanistic insight and molecular basis of cancer and related phenomena. It seeks to promote diverse and integrated areas of molecular biology, cell biology, oncology, and genetics.

Related Collections
With Collections, you can get published faster and increase your visibility.
Metabolic Reprogramming in Cancer
The field of cancer metabolism has expanded rapidly, revealing how metabolic reprogramming drives tumour progression, therapy resistance, and cellular adaptation. Beyond local interactions, accumulating evidence shows that circulating immune cells and blood‑borne factors—including metabolites, cytokines, and extracellular vesicles—actively influence tumour metabolic states. These systemic signals interact with tumour‑resident cells to shape metabolic plasticity, modulate survival pathways, and affect treatment responses.
Within the tumour microenvironment itself, metabolic interplay between cancer cells, immune cells, and stromal components remains a central determinant of tumour behaviour. Altered glycolytic flux, lactate‑driven microenvironmental changes, and cancer stem cell–associated metabolic adaptations contribute to tumour aggressiveness, immune evasion, and resistance mechanisms. Together, these local and systemic dimensions provide an integrated view of how metabolism underpins cancer progression.
This collection supports United Nations SDG 3: Good Health & Well-Being.
Topics of interest include:
- metabolic interactions within the tumour microenvironment
- cancer stem cell metabolism
- glycolytic pathways in tumour promotion
- tumour–immune cell metabolic interplay (local and systemic)
- interplay between circulating blood cells and tumour metabolism
- drug resistance and metabolic shifts
- protein expression in metabolic reprogramming
- role of transcription factors in metabolism
- signal transduction pathways affecting cancer metabolism
Publishing Model: Open Access
Deadline: Dec 31, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in