From Hazard to Asset: How Evolution Turned a Viral Weapon Into a Dietary Tool
Published in Ecology & Evolution and Cell & Molecular Biology

I have always been fascinated by how nature creates endless diversity without altering its core blueprints. While the engine of evolution is propelled by genetic changes hardwired into DNA, RNA editing stands out as a uniquely elegant alternative—it rewrites transcripts on the fly, expanding the functional diversity of the proteome without touching the genome itself [1]. A textbook masterclass of this process occurs in vertebrates through C-to-U editing, driven by the enzyme APOBEC1 (A1). Alongside its essential cofactors (A1CF or RBM47), A1 enters the nucleus on a hyper-specific mission: it binds exclusively to the massive 14-kilobase apoB RNA transcript, and out of all the cytosines available, it targets and edits just one single, precise residue—C6666—to a uracil [2].
Permanently Misnamed: How a Unique Mammalian Trick Defined the Whole APOBEC Family
The way this gene family got its name reveals a fascinating gap between what scientists first discovered and how evolution actually worked. When the first editing enzyme was isolated, researchers named it after its only known function: editing apoB RNA, calling it A1 [3]. As genome sequencing advanced, the family grew to include APOBEC2 through APOBEC5. Herein lies the irony, none of these other family members edit ApoB RNA. In fact, as our study highlights, even A1’s original, ancestral job wasn't to edit RNA at all—it was to target the DNA of retrotransposons. It is a wonderfully funny evolutionary twist that an entire powerhouse family of genomic defenders is now permanently named after a highly specialized, moonlighting job that just one member picked up exclusively in mammals.
What Happened to A1 in Birds? Solving an Evolutionary Cold Case
This remarkable precision is exactly what drew me to a fascinating evolutionary paradox. While the enzyme, its cofactors, and the target transcript are found across almost all vertebrates, this specific editing event has only ever been seen in mammals. Why? This single question launched our genomic investigation. The existing literature offered conflicting clues: some studies suggested certain birds completely lacked A1, while others reported finding it. We wanted to clear the confusion and chart what truly happened to A1 across the avian tree. If it was lost, which bird lineages lost it, and when? If it is still present, what has its real job been in birds all this time?
Initially, we were attempting to untangle a common point of confusion in genomics: gene death is not the same as gene loss. When a gene "dies" via pseudogenization, the sequence remains physically present but non-functional. True gene loss, however, means the physical DNA has been disrupted or completely deleted. While you usually cannot validate this functional impact purely from sequence data, A1 is a rare exception because its very function is editing. This allowed us to hunt for clues in two distinct ways: matching DNA and RNA from the same sample to catch bases modified only in the transcripts, and scanning the genome for the telltale molecular fingerprints A1 leaves directly in the code.
What kept me hooked was the sheer thrill of the chase—investigating gene loss feels remarkably like a high-stakes forensics investigation. As the saying goes, "absence of evidence is not evidence of absence." We cannot simply declare a missing gene lost, because it might just be a case of our inability to find it. Pinpointing when and how this unseen evolutionary event occurred means tracking down the genomic remains of the gene at its proper genetic address (the conserved syntenic location). This requires chasing down disparate clues until independent lines of evidence flawlessly intersect to map out a single ancient event. There is an undeniable rush in watching a chaotic puzzle suddenly snap into focus, elevating a mere educated guess into robust, definitive evidence of evolutionary history.
Lost or Misunderstood? Moving Beyond Conflicting Clues to Define the True Status of Avian APOBEC1
To find out what happened to A1, we analyzed 81 bird species across the entire avian tree. Looking through various genomic datasets, we consistently found a broken gene riddled with mutations. Because these identical flaws appeared across independent data from the same species, we knew it was genuine biology, not a technical glitch. Interestingly, a pattern quickly emerged: 25 of the 33 birds missing the gene were either land fowl (like chickens) or ratites (like ostriches). It was tempting to assume the loss was linked to an obvious shared trait like flightlessness, but biology is rarely that simple. We also found eight other birds from completely unrelated lineages that had lost the gene as well, meaning the real reason behind the loss of A1 is bound to be something much more nuanced.

Figure 1. Inactivating mutations in A1 of birds. (Figure 4 of the article)
Phylogenetic tree of analyzed bird species shown alongside a schematic of the A1 coding region. Branch and taxon colors indicate inferred gene status (black: intact; red: gene loss). Exons 1–5 are represented by horizontal bars (blue: intact; red: deleted; grey: unaligned/missing), with functional residues marked by vertical dashed lines. Small ORF-disrupting mutations, premature stop codons, and repeat insertions are mapped onto their respective positions. Inset alignments provide representative SRA read evidence supporting A1 disruption (for details, see main text).
An Evolutionary Plot Twist: How did a Dangerous Weapon Turn Into a Specialized Tool, and Why?
Seeing A1 present in some birds but missing in others raised a big question: do birds with an intact gene still use it to edit RNA and process fats like mammals do? To find out, we checked the molecular checklist required for this process: the A1 editor, a helper protein (cofactor), and a target RNA with recognition elements. While birds have the helper, their target RNA is riddled with mutations that completely abolish editing. Even worse, the avian A1 protein itself is missing the key amino acids needed to function. Could it be targeting a different RNA instead? We scanned the entire transcriptome of a representative bird, the hooded crow, searching for alternative targets. The results were clear: we found only A-to-I, and not C-to-U editing type, suggesting that the classic mammalian function of A1 might be completely defunct across all birds.

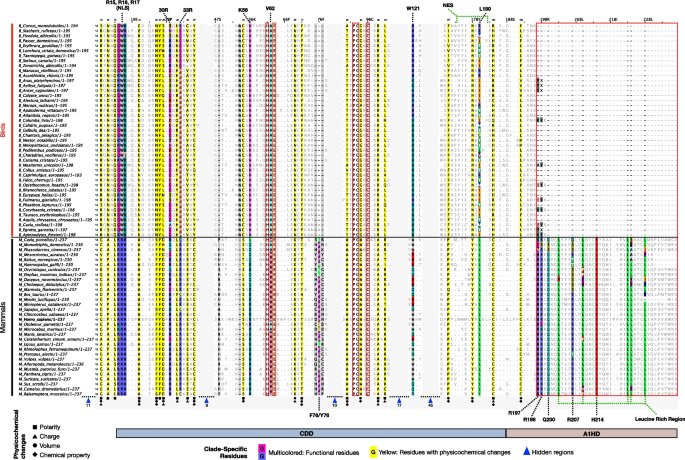
Figure 2. Differences in A1 amino acid composition between birds and mammals (Figure 1 of the article)
Multiple sequence alignment of avian and mammalian A1 proteins, with numbering based on the human sequence. Multicolored columns highlight known functional sites (including RNA editing, NLS, and NES regions), while yellow columns mark bird-mammal substitutions causing predicted physicochemical changes. The red box highlights the mammal-specific C-terminal extension (A1HD region), which is absent or highly reduced in birds, as shown in the domain schematic below (for details, see main text).
If A1 doesn't edit RNA in birds, what is its true purpose? From a dietary standpoint, birds eat much less lipid-rich food than mammals, so they simply don't need a specialized RNA-editing trick to quickly clear fats. Instead, the clues pointed to DNA editing. From lack of a domain in A1, we found that avian A1 cannot form dimers; it remains a monomer. In the molecular world, this makes the enzyme hyper-catalytic—meaning it is highly active but completely blind, losing its specificity and might mutate any cytosine it touches. A hyperactive, non-specific editor loose in the nucleus is a recipe for fatal genomic instability and cancer. So, how do birds survive it? The breakthrough came when we looked at Endogenous Retroviruses (ERVs)—ancient viral "parasites" embedded in the genome. When we scanned bird genomes, we found clusters of mutational scars exactly where these ERVs live. Because ERVs constantly try to replicate and spread, unleashing a hyperactive, non-specific editor on them is the perfect defense strategy, allowing birds to neutralize genomic invaders without needing complex targeting machinery.
Interestingly, this viral-fighting DNA editing function isn't unique to birds; it’s found in early mammals, reptiles, and lizards too. In primates like us, a newer gene family called APOBEC3 took over this job). This suggests that DNA editing—not RNA editing—is actually the ancestral role of A1. But if it’s so beneficial, why are some birds shedding the gene while others hold onto it? The answer lies in the workload. Because birds generally have far fewer ERVs in their genomes than mammals, and thus face a much lower risk of cancer from extensive DNA editing, the evolutionary pressure to keep A1 relaxed. The smoking gun came when we looked at songbirds (Passeroidae, like zebra finches), which recently experienced a massive expansion of new ERV invasions. Sure enough, these exact birds have perfectly intact A1 genes and the highest levels of viral "scars." Ultimately, while A1 is slowly fading out in most bird lineages due to a lack of sufficient viral burden, it remains highly active in the ones facing active genomic invasions.
The Big Picture
When we pieced together our findings with the existing literature, an elegant evolutionary trajectory emerged. A1 did not start as a metabolic tool; it was born as an ancient, "promiscuous" weapon against viral invaders. While birds and reptiles kept this ancestral DNA-defense role, it came with a dangerous trade-off: a loose-cannon DNA editor risked causing cancer. Consequently, whenever viral pressure dropped, avian lineages chose to shed the gene entirely. Mammals, however, took a different path. Once a newer gene family (A3) took over viral defense, A1 was finally freed to evolve. It gained the structural precision needed to bind RNA safely, which ultimately allowed mammals to adapt to fat-rich diets and complex lymphatic systems. It was a stunning example of how a dangerous genomic shield was slowly reshaped into a precise metabolic masterpiece.
References
1. Gommans WM, Mullen SP, Maas S. 2009 RNA editing: A driving force for adaptive evolution? BioEssays 31, 1137–1145. (doi:10.1002/bies.200900045)
2. Davidson NO, Shelness GS. 2000 Apolipoprotein B: mRNA Editing, Lipoprotein Assembly, and Presecretory Degradation. https://doi.org/10.1146/annurev.nutr.20.1.169 20, 169–193. (doi:10.1146/ANNUREV.NUTR.20.1.169)
3. Davidson NO. 1995 Proposed nomenclature for the catalytic subunit of the mammalian apolipoprotein B mRNA editing enzyme: APOBEC-1. National Institutes of Health 1, 3.
Follow the Topic
-
Immunogenetics
Immunogenetics welcomes research in genetics and evolution of the immune system; genetic control of immune response; bioinformatics of the immune system; structure of immunologically important molecules; and immunogenetics of reproductive biology, tissue differentiation, and development.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in