High-density lipoprotein regulates angiogenesis by long non-coding RNA HDRACA
Published in Cell & Molecular Biology
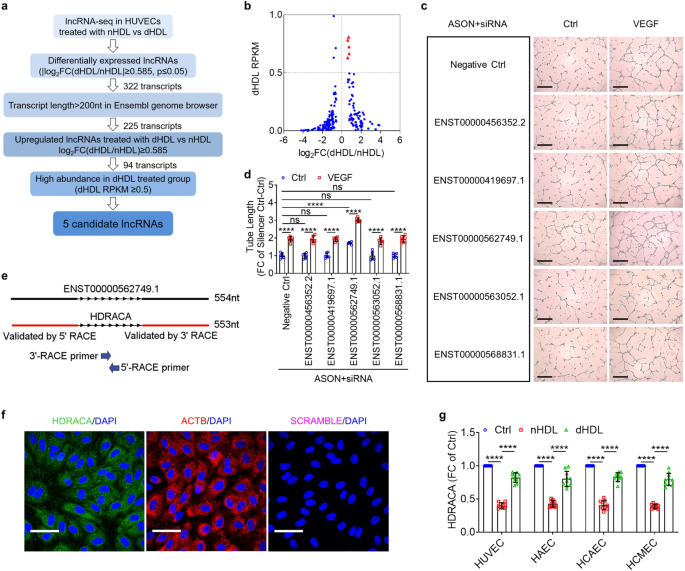
LncRNA HDRACA is involved in the high-density lipoprotein (HDL) regulation of angiogenesis. Normal HDL (nHDL) inhibits the expression of HDRACA to induce angiogenesis, and dysfunctional HDL (dHDL) is much less effective in inhibiting HDRACA expression and inducing angiogenesis.
Currently, no effective methods are available to promote coronary vascular growth in patients with in coronary artery disease (CAD) since myriad risk factors contributing to vascular disease and endothelial dysfunction. A notable and perhaps underestimated risk frequently involves modification of HDL.
HDL is a mixture of multiple proteins, lipids and carrying noncoding RNA. HDL from healthy individuals is called normal HDL (nHDL) and able to induce angiogenesis1, 2. However, the components of HDL are altered or modified in CAD, and HDL becomes dysfunctional1, 3, 4. This dysfunctional HDL (dHDL) is less effective in inducing angiogenesis1, 4. Although several important signaling pathways involved in the regulation of angiogenesis by HDL has been reported1, 2, 5, the mechanisms by which nHDL promotes angiogenesis and dHDL is less effective are not yet fully understood.
Long noncoding RNA (lncRNAs) are arbitrarily defined as a class of ncRNAs comprising more than 200 nucleotides with large quantity and complex functions6. We previously identified lncRNAs that are differentially expressed in endothelial cells after treatment with nHDL or dHDL7. In the present study, by analyzing our previous lncRNA-seq results, we identified a lncRNA that is differentially expressed in endothelial cells stimulated with nHDL or dHDL and is involved in HDL regulation of angiogenesis in CAD. We termed this lncRNA high-density lipoprotein-regulated angiogenesis in coronary artery disease (HDRACA).We showed for the first time that nHDL reduced HDRACA to induce angiogenesis, whereas dHDL from patients with CAD was less able to effectively decrease HDRACA and stimulate angiogenesis.
We found that HDL-bound S1P (sphingosine 1-phosphate) interacted with S1P receptor 1 (S1P1) in the endothelial cells to inhibit HDRACA expression. S1P primarily binds to apolipoprotein M (ApoM) in HDL for transport in the plasma8. S1P in HDL is responsible for many pleiotropic functions of HDL9. The interaction between S1P and S1P1 in the endothelial cells contributes to angiogenesis10.S1P1 is constitutively associated with WW domain-containing E3 ubiquitin protein ligase 2 (WWP2), and WWP2 is able to catalyze Kruppel-like factor 5 (KLF5) ubiquitination11, 12.We found that ApoM-bound S1P induced WWP2 phosphorylation to activate WWP2, which induced the ubiquitination and degradation of Kruppel-like factor 5 (KLF5), a transcription factor that activated HDRACA promoter, leading to decreased transcription of HDRACA in the endothelial cells. Consistent with previous study, our data also showed that dHDL contained lower S1P content than nHDL, which might result from HDL oxidation, abnormal S1P distribution in HDL subfractions or competition between S1P and other ApoM-bound lipids4, 13, 14. Loss of S1P caused dHDL less effective in inhibiting HDRACA expression.
Angiogenesis is a multifaceted and tightly orchestrated process. We investigated the role of HDRACA on endothelial cell cycle, proliferation, migration, tube formation, and apoptosis. Our results showed that HDRACA negatively regulated the cell cycle without affecting endothelial cell apoptosis. Moreover, nHDL promoted cell proliferation and tube formation by downregulating HDRACA in endothelial cells, whereas dHDL had less effect.
In general, lncRNAs participate in transcriptional and post-transcriptional regulation, cell organellar and structural organisation and genome integrity by interacting with other factors15.HDRACA is primarily located in the cytoplasm of endothelial cells. Therefore, it may interact with cytoplasmic fractions to perform its biological functions. Using RNA pull down assays, we found that HDRACA bound to Ras-interacting protein 1 (RAIN), an essential regulatory factor of endothelial cell morphogenesis and blood vessel tubulogenesis16, 17. Our further assays showed that HDRACA hindered the interaction between RAIN and vigilin. As an RNA-binding protein, vigilin can increase or decrease target mRNAs stability by restricting the access of effector proteins to them18. Our data demonstrated that vigilin bound to PCNA mRNA and accelerate PCNA mRNA decay. Moreover, knocking down HDRACA or overexpressing RAIN can interrupt the interaction between vigilin and PCNA mRNA. PCNA can induce different DNA polymerases to bind to DNA at different stages of the cell cycle and promote DNA replication and cell cycle progression19. We found that nHDL increased PCNA expression by downregulating HDRACA to promote endothelial cell proliferation and tube formation, whereas dHDL was less effective. Collectively, nHDL and dHDL differentially mediate angiogenesis by differentially regulating HDRACA expression to affect RAIN-vigilin-PCNA interaction.
HDRACA is poorly conserved across species based on sequence conservation. However, lncRNAs with low sequence conservation can be conserved at other levels, including secondary or high-order structures, short motifs for protein binding, and positions and functions of lncRNAs20. Human and mouse RAIN were highly conserved. Therefore, we hypothesized that human HDRACA bound to mouse RAIN and exerted a similar regulatory effect on angiogenesis. When we ectopically expressed human HDRACA in mouse endothelial cells, we found that HDRACA also inhibited the proliferation and tube formation of mouse endothelial cells. In addition, human HDRACA can also bind to mouse RAIN. Further assays that ectopically expressing human HDRACA in C57/BL6 mice confirmed the inhibitory effect of HDRACA on angiogenesis in vivo.
Arteriosclerosis obliterans (ASO) and CAD are manifestations of atherosclerosis with similar risk factors and metabolic disorders. The endothelial cells of the lower limb arteries in patients with ASO showed upregulation of KLF5 and HDRACA, and downregulation of PCNA. Furthermore, the S1P content in HDL was inversely correlated with HDRACA levels in the endothelial cells of the lower limb arteries. This finding preliminarily confirmed the change in S1P-KLF5-HDRACA-PCNA signaling in ASO, and indicated the potential translational relevance of HDRACA.
In summary, our study reveals a novel HDRACA dependent mechanism by which nHDL induces angiogenesis and explain why dHDL does not induce angiogenesis. HDRACA can serve as a novel therapeutic target for angiogenesis in patients with CAD.
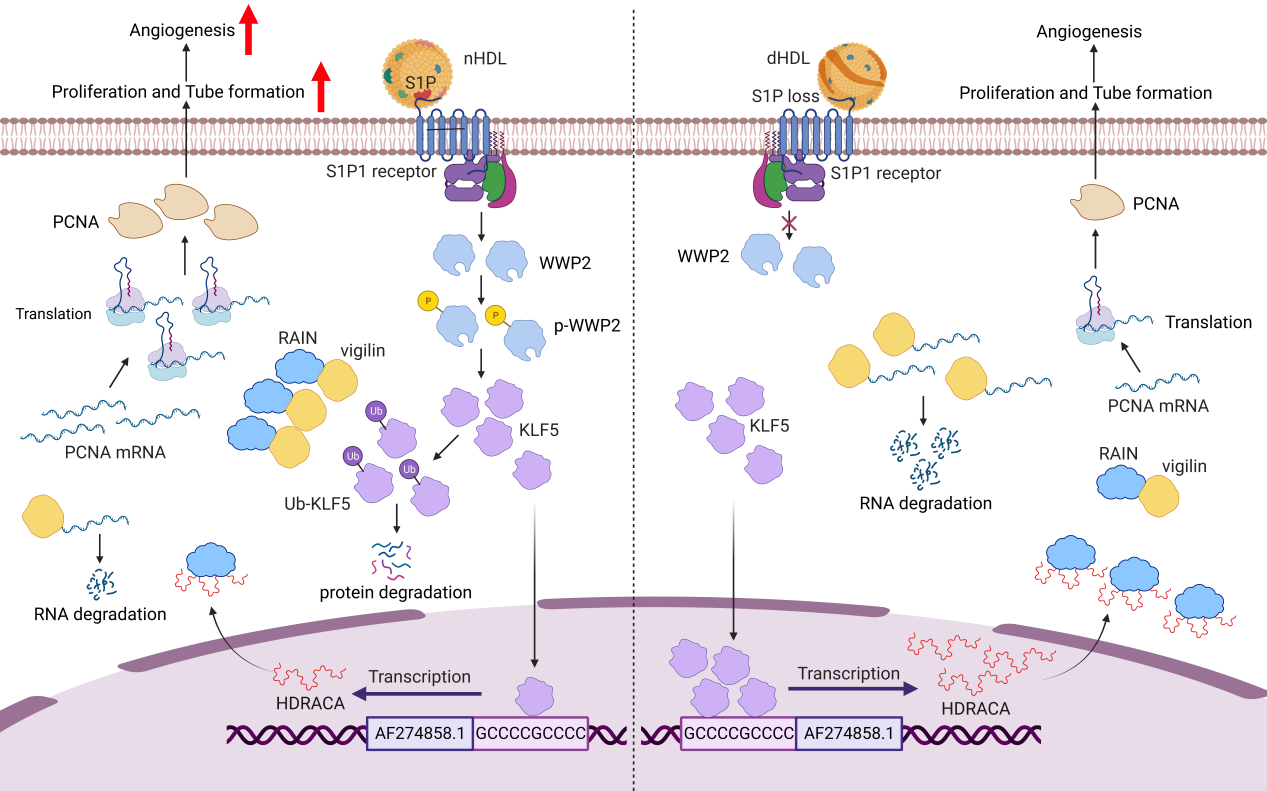
Figure. Graphical illustration of HDL-HDRACA regulatory mechanism for angiogenesis.
https://doi.10.1038/s41392-023-01558-6
Reference
- Li, H.M. et al. Angiogenic and Antiangiogenic mechanisms of high density lipoprotein from healthy subjects and coronary artery diseases patients. Redox Biol.36, 101642 (2020).
- Jin, F. et al. High-density lipoprotein (HDL) promotes angiogenesis via S1P3-dependent VEGFR2 activation. Angiogenesis.21, 381-394 (2018).
- Rosenson, R.S. et al. Dysfunctional HDL and atherosclerotic cardiovascular disease. Nat Rev Cardiol.13, 48-60 (2016).
- Sattler, K. et al. Defects of High-Density Lipoproteins in Coronary Artery Disease Caused by Low Sphingosine-1-Phosphate Content: Correction by Sphingosine-1-Phosphate-Loading. J Am Coll Cardiol.66, 1470-1485 (2015).
- Tan, J.T. et al. High-density lipoproteins augment hypoxia-induced angiogenesis via regulation of post-translational modulation of hypoxia-inducible factor 1α. Faseb j.28, 206-217 (2014).
- Liu, L. et al. Noncoding RNAs: the shot callers in tumor immune escape. Signal Transduct Target Ther.5, 102 (2020).
- Liu, X. et al. High density lipoprotein from coronary artery disease patients caused abnormal expression of long non-coding RNAs in vascular endothelial cells. Biochem Biophys Res Commun.487, 552-559 (2017).
- Christoffersen, C. et al. Endothelium-protective sphingosine-1-phosphate provided by HDL-associated apolipoprotein M. Proc Natl Acad Sci U S A.108, 9613-9618 (2011).
- Sattler, K. & Levkau, B. Sphingosine-1-phosphate as a mediator of high-density lipoprotein effects in cardiovascular protection. Cardiovasc Res.82, 201-211 (2009).
- Tawa, H. et al. Role of afadin in vascular endothelial growth factor- and sphingosine 1-phosphate-induced angiogenesis. Circ Res.106, 1731-1742 (2010).
- Oo, M.L. et al. Engagement of S1P₁-degradative mechanisms leads to vascular leak in mice. J Clin Invest.121, 2290-2300 (2011).
- Fu, J. et al. WWP2 Promotes Odontoblastic Differentiation by Monoubiquitinating KLF5. J Dent Res.100, 432-439 (2021).
- Sattler, K.J. et al. Sphingosine 1-phosphate levels in plasma and HDL are altered in coronary artery disease. Basic Res Cardiol.105, 821-832 (2010).
- Sevvana, M. et al. Serendipitous fatty acid binding reveals the structural determinants for ligand recognition in apolipoprotein M. J Mol Biol.393, 920-936 (2009).
- Oo, J.A., Brandes, R.P. & Leisegang, M.S. Long non-coding RNAs: novel regulators of cellular physiology and function. Pflugers Arch.474, 191-204 (2022).
- Barry, D.M. et al. Rasip1-Mediated Rho GTPase Signaling Regulates Blood Vessel Tubulogenesis via Nonmuscle Myosin II. Circ Res.119, 810-826 (2016).
- Koo, Y. et al. Rasip1 is essential to blood vessel stability and angiogenic blood vessel growth. Angiogenesis.19, 173-190 (2016).
- Cheng, M.H. & Jansen, R.P. A jack of all trades: the RNA-binding protein vigilin. Wiley Interdiscip Rev RNA.8, e1448 (2017).
- O'Donnell, M., Langston, L. & Stillman, B. Principles and concepts of DNA replication in bacteria, archaea, and eukarya. Cold Spring Harb Perspect Biol.5 (2013).
- Diederichs, S. The four dimensions of noncoding RNA conservation. Trends Genet.30, 121-123 (2014).
Follow the Topic
-
Signal Transduction and Targeted Therapy
This is an international, peer-reviewed, open-access journal publishing articles related to signal transduction in physiological and pathological processes, alongside signal transduction-targeted therapeutics in the form of biological agents and small molecular drugs used to treat human diseases.

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in