The Return of the Builder: Cancer as a Misguided Architect
Published in Cancer and Cell & Molecular Biology
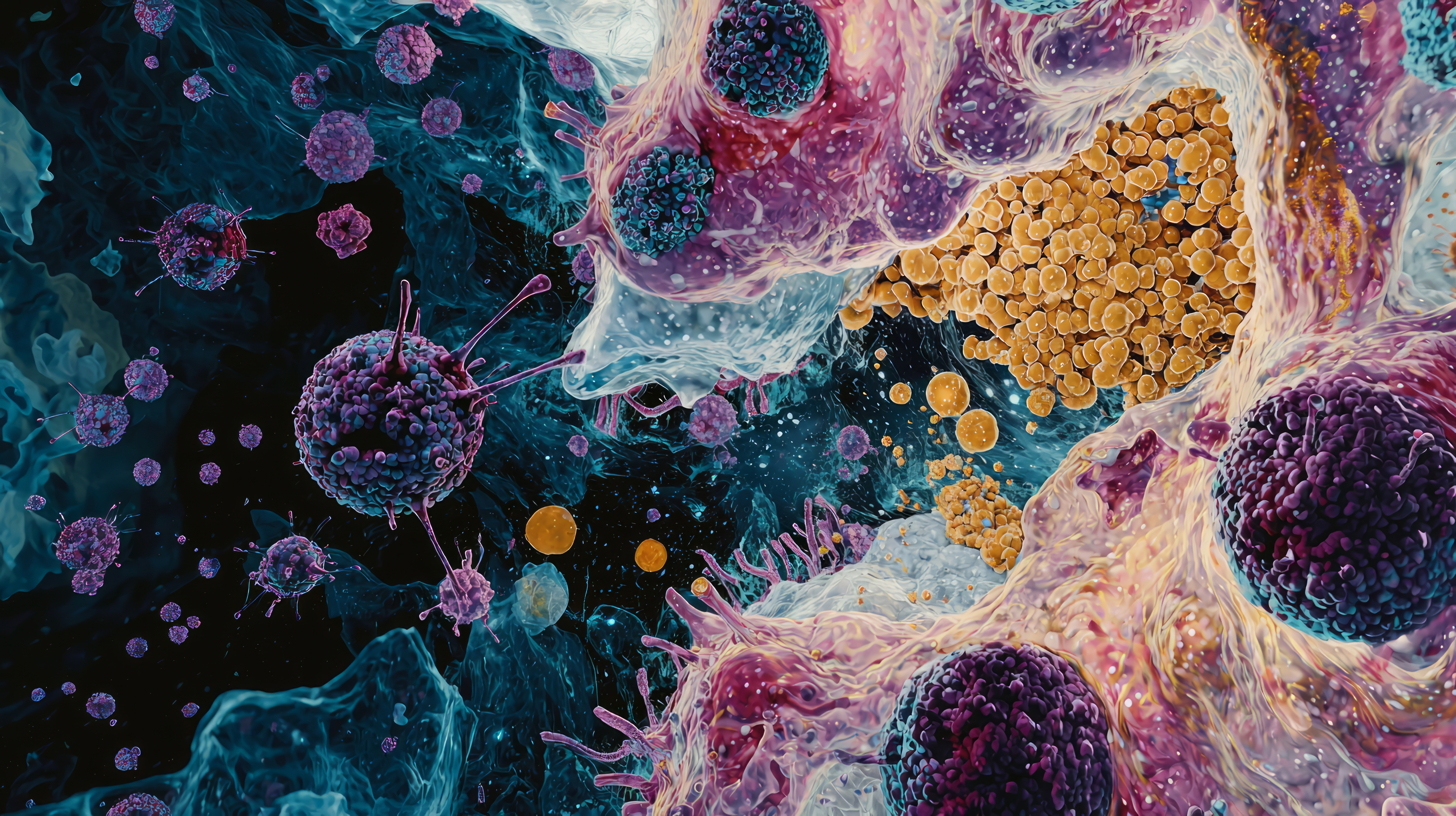
When I first observed a tumor under the microscope, I didn’t see random disarray.
Instead, I saw something strangely intentional. The cells were densely packed, arranged in whorls and layers, forming ducts, clefts, and gland-like spaces that echoed the structure of normal organs. There was a surprising symmetry to the chaos. What I saw looked less like biological collapse and more like mimicry—a dark imitation of design.
That resemblance, I would later learn, was not coincidental. It was a replay of developmental blueprints—the reactivation of instructions that are meant to operate only once, and only in a very different context.
For more than a century, pathologists have noticed this uncanny similarity. In 1907, Karl Landsteiner, the Nobel Prize–winning Austrian immunologist, described neuroblastomas as "embryonal" tumors. Others noted how Wilms tumors of the kidney retained nephrogenic rests—embryonic remnants that failed to mature. Teratomas, some of the strangest tumors in pathology, contained teeth, bone, hair, even retinal structures. These weren’t the products of mutations alone, but of developmental machinery run amok.
As cancer biology advanced in the late 20th and early 21st centuries, molecular studies began to confirm what pathologists had long suspected. Tumors weren’t just growing uncontrollably—they were following specific, familiar programs. Genes involved in embryonic development were being reexpressed in cancer. Pathways such as Hedgehog, Wnt, Notch, and TGF-β—critical for organogenesis and morphogenesis in the embryo—were being hijacked by tumor cells to promote survival, proliferation, invasion, and metastasis.
For example, branching morphogenesis, a process essential to the development of the lung, kidney, and mammary gland, has parallels in invasive breast cancers. The very genes that guide epithelial cells to form complex ductal networks during embryogenesis—such as FGF10 and its downstream effectors—are co-opted in malignancy to carve paths through tissue.
The parallels go beyond molecular signatures. Development and cancer both rely on similar cellular behaviors: rapid proliferation, epithelial-to-mesenchymal transition (EMT), angiogenesis, and tissue remodeling. During development, these processes are tightly regulated and coordinated. In cancer, they are deregulated but not directionless. Tumor cells often engage in a form of pathological choreography, mobilizing as if toward organogenesis, but with no goal other than expansion.
This has led to a profound shift in how we interpret cancer. Rather than viewing tumors as anarchic, many scientists now see them as organisms within organisms—structures that co-opt developmental processes for pathological ends. The resemblance to embryonic tissues isn’t superficial; it is foundational.
This developmental framing has given rise to an entire subfield of oncology that investigates the shared principles of development and cancer. Organoid technology, in which stem cells or tumor cells are cultured into 3D mini-organs, has become a powerful tool for studying how tissue architecture influences cell fate. Organoids derived from patient tumors often retain features of their parent tissue, including differentiation potential and responsiveness to signals. These models allow researchers to test how developmental cues—like gradients of morphogens or mechanical forces—affect tumor growth.
Cancer stem cells further complicate the picture. These rare, plastic subpopulations within tumors exhibit self-renewal and multipotency, much like embryonic stem cells. They are thought to contribute to therapy resistance and disease relapse. Understanding how they maintain their stem-like properties has led researchers back to early developmental regulators. Transcription factors such as SOX2, OCT4, and NANOG—mainstays of the embryonic gene expression landscape—are often expressed in aggressive cancers.
Clinically, this understanding is opening new doors. Therapies targeting embryonic pathways are being evaluated in clinical trials. Inhibitors of the Hedgehog pathway have been approved for basal cell carcinoma and are being tested in medulloblastoma. Wnt pathway inhibitors are under study for colorectal and triple-negative breast cancers. The goal is not to suppress growth indiscriminately, but to selectively interrupt the signals that tumors have borrowed from their developmental past.
But the implications are not purely therapeutic. They are conceptual. If tumors are misdirected versions of our developmental selves, then cancer is not just an error in coding or regulation. It is a misapplication of memory—a script that was supposed to be read once, cleanly, and then closed. Cancer reopens that script, improvising from its lines but distorting the plot.
This changes how we talk about cancer. It shifts the metaphor. Cancer is not chaos. It is order in the wrong place. It is not a fall from structure, but a structure bent toward the wrong purpose. The embryo in the mirror is not poetic flourish—it is the biological reality of cells remembering what they were once asked to do.
And that memory is powerful. It explains how tumors self-organize, how they resist, how they adapt. It also gives us a roadmap. Because the same pathways that build can be unbuilt. The same logic that constructs can be rewired.
To understand cancer fully, we must look not only at what cells have lost—but at what they have reawakened. The embryo is not merely a metaphor for the tumor. It is its ancestor.
And in that ancestry, we may find both the origins of malignancy and the clues to its defeat.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in
Great observations by you of a sort of "Call of the Wild" concept of Jack London of a civilized dog reverting to a primal being after being physically abused. Cells abused by radiation, toxic chemicals, and the damages from aging get their civilizing mechanisms damaged and stripped away to reactivate their primitive nature embodied in the embryonic state, which replays the evolutionary developmental history of the last 500 million years or so of multicell organisms. I am working on an aspect of a possible tubing-cell paper, that proposes that in the beginning over 500 million years ago, a new cell type like a neuron cell with a long filament, or a fungus with a long filament, started to coil its long filament to form a coiled tube to provide a larger fluid transporting tube, as well as to provide stiffness to form primitive backbone structures. See my website: Microscope Images | Golf Science Guru of a plant leaf using tubing-cells. I wonder if you have seen any striated tubing-cells in cancel images. If you saw any, please let me know. Thank you.