Behind the Paper: Reverse Engineering Morphogenesis
Published in Physics, Protocols & Methods, and Cell & Molecular Biology
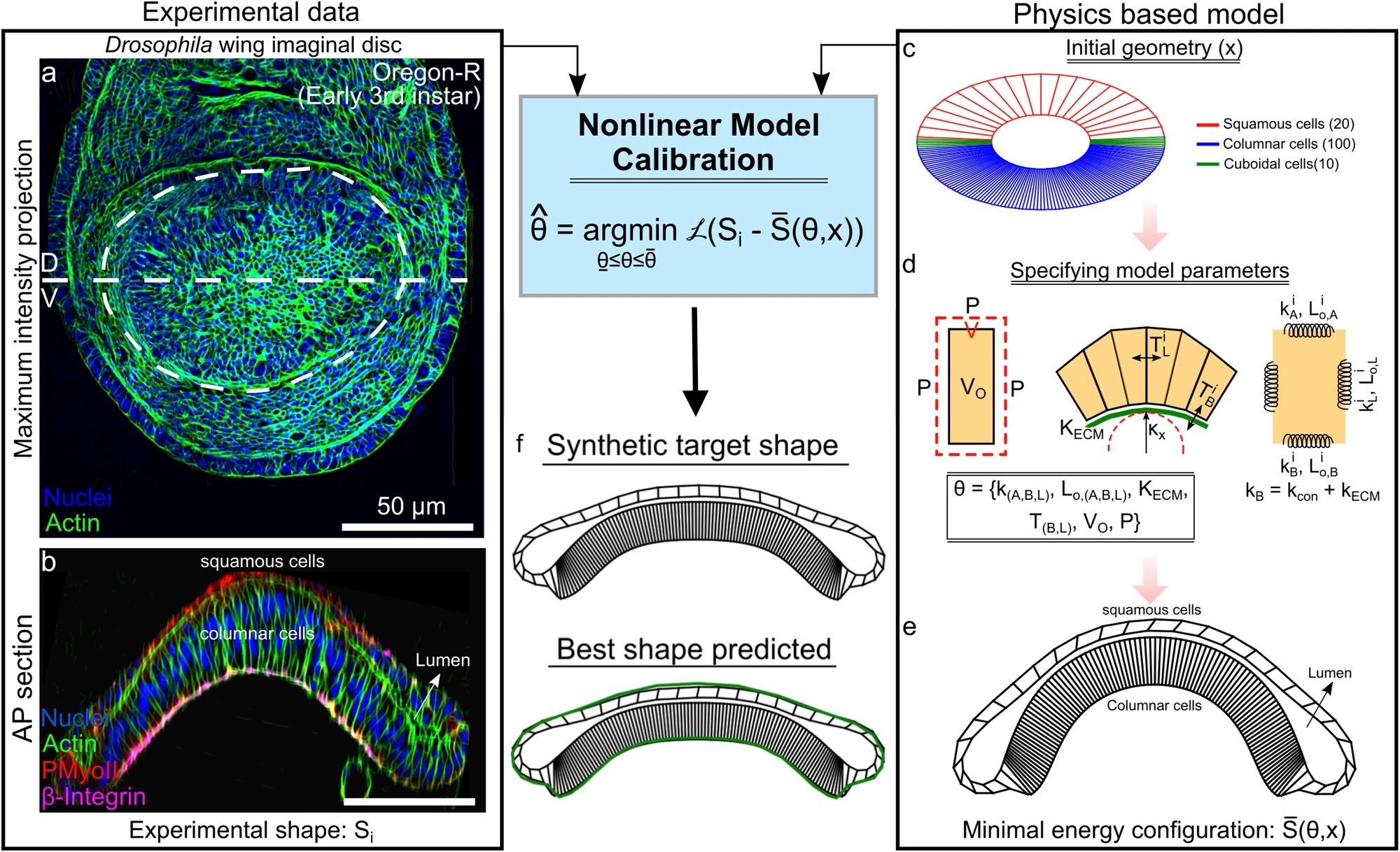
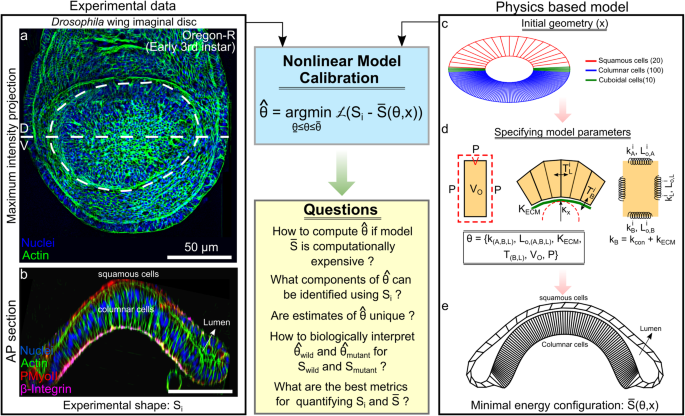
Computational models are essential tools for elucidating the mechanisms of biological processes. However, due to the complexity of biological systems, these models can take days to evaluate on modern computers. Additionally, model parameters must be calibrated, i.e., fine-tuned, using experimental data. Our recent paper proposes a new hybrid approach to optimally estimate parameters in an expensive computer model that predicts the shape of Drosophila wing imaginal discs1. To reduce computational load, we created a Bayesian optimization (BO) framework that uses a machine learning-based surrogate model2. This approach utilized a physical based model using the software Surface Evolver that we incrementally adjust to fit experimental data, outputting parameters that are difficult to measure experimentally3. We can use these parameters to predict mechanisms for morphological changes, effectively reverse engineering morphogenesis of the wing disc.
The wing imaginal disc is a fluid-filled sac surrounded by epithelial cells, which are enclosed by a thin extracellular matrix (ECM). The biophysical model simplifies the shape of the wing disc as defined by physical forces between each cell and each other and each cell and the ECM. The model defines the cellular forces in a set of energy functions to reflect the contractility of the cell, a key factor in cellular shape4. We described each cell edge as a Hookean spring with a natural length. Higher cell contractility increases stiffness, meaning cells are more resistant to size changes; Hooke’s law is used to quantify this resistance to change as cellular energy. Likewise, energy functions were defined for the volume of the cell, cellular adhesion to the ECM, and the ECM itself (modeled as an elastic string). Once the Surface Evolver environment is set up to our initial specifications, we run iterations to minimize the system’s total energy3. Over time, the model converges to the lowest energy configuration that closely matches the observed shape of the wing disc cross section.
To better calibrate the model to the experimental data and quantify the error, we utilized a BO-based computational pipeline. We tested metrics for measuring the similarity of the experimental and model wing disc shapes, identifying Fréchet distance, a metric describing the closeness of two curves, as the best at quantifying similarity. In brief, the BO framework uses iterations of simulated wing disc shapes, minimizing the Fréchet distance at certain points, training the model to fit experimental data. After enough iterations, we recover a set of parameters that describe the shape of the wing disc, specifically cellular contractility. These parameters are especially useful, as they are not obtainable through experimental measurements.
To verify the BO framework, we tested the model with an imaginal disc that had undergone degradation of the ECM. Experimentally, ECM degradation from a pharmaceutical agent produces an inverted disc5; our model fits this new shape and predicts a decrease in contractility across the disc . Our BO framework revealed that degradation of ECM results in a global increase in contractility, most significantly on the basal side of the wing disc. Experimentally, we measured phosphorylated non-muscle myosin (pMyoII), a regulator of contractile forces in a tissue, and observed a decrease in levels on the basal side, verifying that the disc increases basal contractility to sustain the bent shape of the wing disc6. These results validate our model with the BO framework as an accurate predictor of morphological changes.
Next, we used the BO framework to predict mechanisms for morphological changes of specific genetic perturbations. We used a genetic variant that allowed us to knockdown or over-express the mechanosensitive membrane protein Piezo. Our model was fit to mutant wing discs and predicted a decrease of contractility with Piezo knockdown and an increase in contractility with over-expression. These results were again verified experimentally with pMyoII measurements. Our model also predicted a decrease in ECM contractility with Piezo over-expression, which aligns with a previous study that observed the degradation of the ECM through Piezo in Drosophila midgut7. Taking other studies into consideration, we hypothesize that Piezo regulates the protein Rho1, which is upstream of pMyoII and may affect pMyoII levels, leading to the predicted changes in contractility 7,8. Future work would need to be done to confirm the exact mechanism of Piezo’s impact on contractility. Our work highlights the importance of Piezo on Drosophila development, opening further areas for research with Drosophila. Just as impactful, though, is our demonstration of Bayesian Optimization as an effective strategy for modeling complex biological processes. Our BO framework can successfully predict experimental data using only an image of a wing disc cross-section. Future work is needed to extend BO to further define morphogenesis as an optimization problem between experimental data and model outputs. Moreover, this strategy could be applied to other fields, such as embryogenesis, plant development, or cancer mechanics. As this BO strategy does not rely on a specific modeling software, it could be applied to almost any computational model, making it a powerful tool for understanding complex physical systems.
References
- Kumar N, Mim MS, Dowling A, Zartman JJ. Reverse engineering morphogenesis through Bayesian optimization of physics-based models. Npj Syst Biol Appl. 2024;10(1):1-14. doi:10.1038/s41540-024-00375-z
- Befort BJ, DeFever RS, Tow GM, Dowling AW, Maginn EJ. Machine Learning Directed Optimization of Classical Molecular Modeling Force Fields. J Chem Inf Model. 2021;61(9):4400-4414. doi:10.1021/acs.jcim.1c00448
- Brakke KA. The Surface Evolver. Exp Math. 1992;1(2):141-165. doi:10.1080/10586458.1992.10504253
- Guo H, Swan M, He B. Optogenetic inhibition of actomyosin reveals mechanical bistability of the mesoderm epithelium during Drosophila mesoderm invagination. Bagnat M, Banerjee U, Streichan SJ, Suzanne M, eds. eLife. 2022;11:e69082. doi:10.7554/eLife.69082
- Nematbakhsh A, Levis M, Kumar N, Chen W, Zartman JJ, Alber M. Epithelial organ shape is generated by patterned actomyosin contractility and maintained by the extracellular matrix. PLOS Comput Biol. 2020;16(8):e1008105. doi:10.1371/journal.pcbi.1008105
- Matsumura F, Ono S, Yamakita Y, Totsukawa G, Yamashiro S. Specific Localization of Serine 19 Phosphorylated Myosin II during Cell Locomotion and Mitosis of Cultured Cells. J Cell Biol. 1998;140(1):119-129.
- Lee J, Cabrera AJH, Nguyen CMT, Kwon YV. Dissemination of RasV12-transformed cells requires the mechanosensitive channel Piezo. Nat Commun. 2020;11(1):3568. doi:10.1038/s41467-020-17341-y
- Pardo-Pastor C, Rubio-Moscardo F, Vogel-González M, et al. Piezo2 channel regulates RhoA and actin cytoskeleton to promote cell mechanobiological responses. Proc Natl Acad Sci U S A. 2018;115(8):1925-1930. doi:10.1073/pnas.1718177115
Follow the Topic
-
npj Systems Biology and Applications
An online Open Access journal dedicated to publishing the premier research that takes a systems-oriented approach and encourages studies that integrate, or aid the integration of, data, analyses and insight from molecules to organisms and broader systems.

Related Collections
With Collections, you can get published faster and increase your visibility.
Mechanistic and data-driven modeling in mathematical oncology
Publishing Model: Open Access
Deadline: Dec 02, 2026
Systems mechanobiology
Publishing Model: Open Access
Deadline: Sep 28, 2026




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in