Big box, small box: bacteria either have all around big niches or all around small niches.
Published in Ecology & Evolution
The world is made up of a bunch of different environmental conditions, like how much water is available, how acidic the soil is, how hot it is, etc. To live and reproduce, species need to successfully navigate all these different environmental conditions. How many kinds of environmental conditions species can exist in determines their “niche.” Species that can exist in a wide range of environmental conditions are “generalists” and those that only exist in a narrow range are “specialists.” Understanding whether species are generalists or specialists should consider their ability to exist across all these different environmental conditions (i.e., multidimensionality).
We looked at the multidimensional niches of >1200 prokaryotes across the continental United States. This was made possible by the recent launch of the National Science Foundation's flagship repository of extensive environmental and ecological data, National Ecological Observatory Network (NEON). With NEON data, we assessed whether generalization and specialization are multidimensional processes in which species either only generalize (i.e., multidimensional generalization) or only specialize (i.e., multidimensional specialization) on all niche dimensions.
You might be asking “Why should we care if multidimensional specialization and generalization is common?” Which is a fair question as it does seem abstract. Talking about dimensions brings up Sci-Fi-esque feelings and that's without bringing up the industry term for multidimensionality – hypervolume – that has always reminded me more of Star Trek than actual science. But the reason why this matters is that, if multidimensional specialization and generalization exist, then the environmental conditions that species can withstand increase and decrease exponentially. For example, if we just look at three different dimensions (e.g., water, temperature, and pH), multidimensional generalization increases the kinds of suitable habitat to the power of three (i.e., the number of dimensions)! Whereas multidimensional specialization decreases suitable habitat by the power of 1/3 (i.e., cubed). That has massive implications for what kinds of habitats species would find suitable with multidimensional generalists being resilient to environmental change and multidimensional specialists being very sensitive since any slight deviation in any one environmental condition would make a habitat unsuitable. So, we asked: 1) how common is multidimensional generalization and specialization and then 2) what are the implications of multidimensional specialization and generalization for prokaryotic evolution, community composition, and community structure.
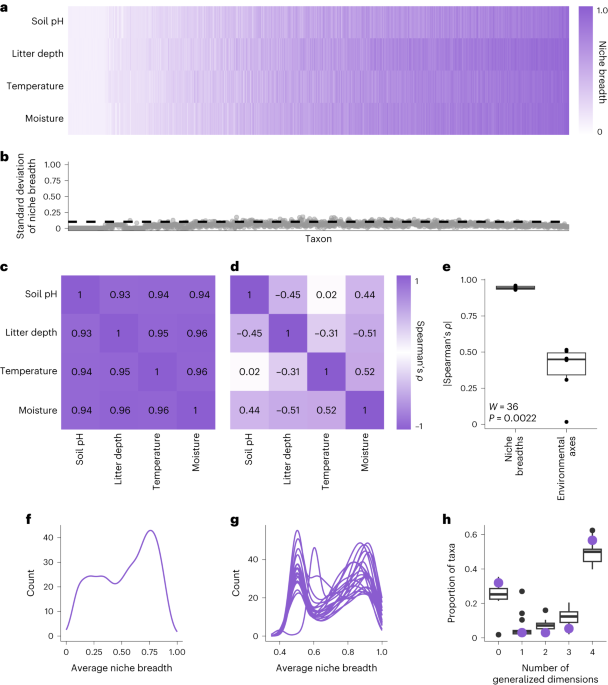
We found that ~90% of prokaryotes are in fact either multidimensional generalists or multidimensional specialists (Figure 1a). This indicates that multidimensional generalization and specialization are almost universal, opposing niche trajectories in soil prokaryotes where specialization (or generalization) on one dimension has consequences for other dimensions. Multidimensional specialization and generalization as opposing trajectories is further supported by how multidimensional specialization and generalization evolve (Figure 1b). We found that multidimensional specialization and generalization are highly conserved across prokaryotes with transitions from generalist to specialist (and vice versa) occurring 90% less often than random chance. This then raises an interesting question about why these two niche trajectories are conserved. We think that multidimensional generalist and specialist lifestyles are conserved because ecological strategies to specialize (or generalize) in one niche dimension may have consequences for specialization and generalization in all other important niche dimensions. Consequently, our finding that multidimensional generalization and specialization are ubiquitous, mutually exclusive, and conserved features of prokaryotic ecology opens new lines of research. For example, are ecological strategies for generalization and specialization incompatible so that, if selection forces a species to specialize on one dimension, a species has a harder time following generalist strategies in other dimensions (e.g., if a microbe has to make spores to specialize in drought conditions, that spore-forming strategy makes it harder to be a generalist in other dimensions like temperature).

We then asked what these conserved, opposing niche trajectories mean for how species are organized in their communities. First, we found that multidimensional generalists dominate their communities by being 73-times more abundant than multidimensional specialists (Figure 2a). The dominance of multidimensional generalists is likely to increase resilience of microbiomes because the organisms that have the most flexible niche requirements – the multidimensional generalists – make up most of their communities. So, if there is a change to their environment, this means that communities predominantly consist of microbes that have leeway to deal with these environmental changes. However, when we looked at how these communities are structured, we found that multidimensional specialists are almost twice as structurally important on average than multidimensional generalists (Figure 2c). Specifically, these microbes associate with nearly double the number of other microbes in the network of community interactions, and these microbes are known to provide important community services, like detoxification and nutrient cycling, that structure community dynamics and ecosystem services (Figure 2d). This suggests that, while a lot of the community may be flexible in terms of environmental conditions (i.e., the multidimensional generalists), the microbes that define and structure these communities are more likely to be extra sensitive to environmental disturbances because their niches are constrained in multiple dimensions. So, most of the community may be very flexible, but the structurally important microbes may be especially sensitive. Does this mean that the many benefits microbiomes provide are sensitive or resilient to changing environmental conditions? Future work understanding what matters more for community stability will be important, such as asking: is community stability mostly impacted by the dominance of multidimensional generalists, the services provided by specialists, or does it depend on whether the specific ecosystem relies on generalist or specialist services?

Putting it all together, generalization and specialization are not only common in soil prokaryotes, but they are also mutually exclusive processes, suggesting that specializing or generalizing on any one dimension means developing ecological strategies that make it hard to do the opposite on other dimensions. Further, these opposing niche trajectories also shape 1) how prokaryotes evolve with multidimensional specialization and generalization seemingly locking species into one of these two trajectories and 2) how communities are structured with microbiomes being dominated by multidimensional generalists while multidimensional specialists do the heavy lifting of structuring communities.
Follow the Topic
-
Nature Ecology & Evolution
This journal is interested in the full spectrum of ecological and evolutionary biology, encompassing approaches at the molecular, organismal, population, community and ecosystem levels, as well as relevant parts of the social sciences.

Related Collections
With Collections, you can get published faster and increase your visibility.
Biodiversity and ecosystem functioning of global peatlands
Publishing Model: Hybrid
Deadline: Jul 27, 2026
Understanding species redistributions under global climate change
Publishing Model: Hybrid
Deadline: Dec 31, 2026





Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in