Extracellular rRNA provides a window on taxon-specific microbial cell lysis
Published in Earth & Environment

More than 30 years ago, three papers were published in Nature1,2,3 showing the enormous abundance of viruses in seawater and their role as mortality agents of heterotrophic bacteria and phytoplankton (Fig. 1); thus, setting the stage for a new paradigm in biological oceanography, and more broadly, for the role of viruses in ecosystems. Since then, thousands of papers have been published on viruses in the sea, documenting their abundance, diversity and ecosystem functions. Yet, despite widely accepted estimates of virus-mediated microbial mortality, indicating that viruses kill about 20 to 30% of the living biomass in the oceans each day4,5, these are bulk estimates that do not provide information on how lysis is distributed among thousands of microbial taxa. Identifying the taxa undergoing lysis has been one of the most important outstanding questions since viruses were recognized as being major agents of microbial mortality. We have tackled this problem by establishing the use of extracellular ribosomal RNA (rRNAext) to estimate taxon-specific cell lysis of microbes.
 Nat Rev Microbiol 5, 801-812; https://doi.org/10.1038/nrmicro1750.")
Figure 1 Occurrence of microbes and viruses in seawater, observed using epifluorescence microscopy. Image source: Suttle (2007) Nat Rev Microbiol 5, 801-812; https://doi.org/10.1038/nrmicro1750.
The rRNAext is the result of cell lysis
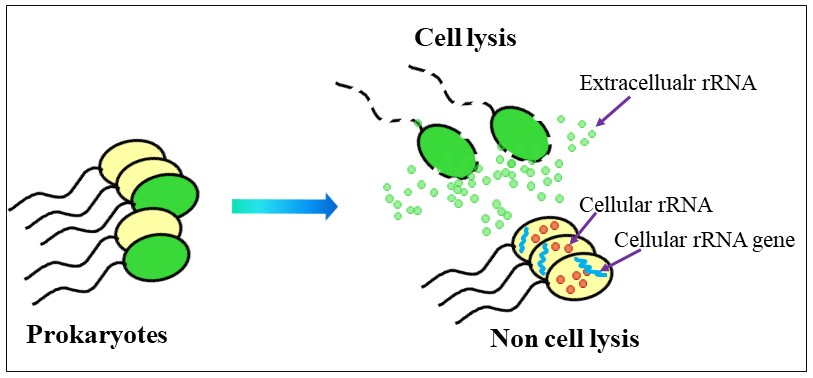
For years, we were surprised by large amounts of ribosomal RNA (rRNA) “contamination” that was present in 0.22-µm filtered seawater collected to investigate the diversity of RNA viruses. We hypothesized that rRNA was released by cell lysis (Fig. 2), and that this rRNAext could be sequenced to reveal the taxonomic composition of the cells that had died.
.")
Preliminary investigations using quantitative reverse-transcription PCR (qRT-PCR) revealed that there were typically millions of copies of rRNAext in each mL of 0.22-µm-filtered seawater, and that it was stable for days when incubated on the benchtop. This indicated that the rRNAext was likely in the form of ribosome complexes that prevent degradation of the RNA, consistent with observations that rRNA complexed with ribonucleoproteins is protected from nucleases6,7.
To confirm the origin of rRNAext, we conducted laboratory experiments with marine bacteria, their viruses and grazers. We showed that rRNAext is produced when bacteria are infected by viruses, and that grazing by protists does not contribute to rRNAext production; in fact, protistan grazing was associated with a decrease in rRNAext. Given that viral lysis accounts for approximately half of the mortality of prokaryotes in seawater2,8,9 and kills approximately 20 to 30% of the standing stock of bacteria each day4,5, viral lysis is likely responsible for most of the rRNAext production. However, other causes of cell lysis such as programmed cell death10, predation by Bdellovibrio and like organisms (BALOs)11, and bacteriocins12,13, may also contribute to the production of rRNAext, although there is no evidence that these processes are quantitatively significant relative to viral lysis.
. The red dot indicates ribosomes.")
rRNAext provides a window on taxon-specific microbial cell lysis
In addition to the rRNAext, we also sequenced the cellular rRNA genes (rRNAgenecell) and cellular rRNA (rRNAcell) (Fig. 3). The taxa in the rRNAext pool reflect cell lysis, while taxa in the rRNAgenecell and rRNAcell pools indicate taxa for which intact cells are present in the water. By comparing the taxa that are detected in each of the three pools (rRNAgenecell, rRNAcell, and rRNAext), we can distinguish among taxa undergoing lysis (ongoing-lysis or recent-lysis), taxa in which lysis has already occurred (prior-lysis), and cells in which lysis has not occurred (no-lysis). MoRS allowed us to
- detect lysis in thousands of different prokaryotic taxa
- show that most lysis occurred prior to the time of sampling
- detect lysis in up to 34% of the taxa detected in coastal seawater samples
- detect lysis in 31 out of 36 prokaryotic phyla, including 19 phyla from which viruses have not been reported, indicating that lysis is widespread across prokaryotic phyla
- show that the taxa in which lysis occurred varied greatly among seawater samples (Fig. 4).
- show that of the taxa in which lysis was detectable, only a maximum of 8% were present in the cellular fraction, indicating that in most cases lysis occurred prior to sampling
- show that lysis was undetectable in most taxa at the time of sampling
 represents a ribosomal ASV, while the size indicates its relative abundance; The line (edge) links ASVs to the seawater samples (square node) in which they were detected. Image source: Zhong et al., (2022) ISME J; https://doi.org/10.1038/s41396-022-01327-3.")
Cell lysis is uneven among microbial taxa
Sequencing of the rRNAext reveals the taxa in which lysis has occurred; if the rRNAext is corrected for differences in “rRNA copy number per cell” that occur among taxa and with physiological state, the ratio of free rRNA (rRNAext) to cellular rRNA (rRNAcell) provides an index of the relative amount of lysis in each taxon. Thus, a cell-lysis index (CLI) can be calculated for individual taxa from the ratio of the relative abundance of rRNAext to rRNAcell. Our results revealed ongoing lysis in 128 taxa, with the CLI varying across depths and time. The results also showed that the CLI is highly uneven among taxa within a seawater sample. This is important, as most current models of microbial mortality assume lysis is evenly distributed among taxa14.
Rare species are associated with high lysis while dominant taxa are associated with low lysis.
An enduring puzzle in marine microbial ecology is the relationship between viral infection and the structure of prokaryotic communities, and in particular the relationship between the abundance of specific taxa and viral lysis. It has been proposed that lysis rates are relatively low for cells in the most abundant taxa, and higher in relatively rare bacteria that are capable of fast growth15. This view is congruent with “Kill the Winner”, a model, in which viral lysis prevents the most ecologically “fit” cells from dominating the community16,17. Conversely, cells associated with rare taxa may be subject to significant viral lysis18. Here, by using MoRS to estimate taxon-specific cell lysis, we demonstrated that high lysis was associated with taxa that were relatively rare (such as copiotrophs), while low lysis was often associated with taxa that were abundant, such as those in the SAR 11 group. Therefore, it is likely that rare species are the result of high lysis, whereas dominants are likely to be successful in part due to relatively low rates of lysis.
In summary, MoRS offers a powerful new approach for understanding how taxon-specific lysis shapes microbial communities and processes, and provides an important tool in our efforts to explain the distribution and abundance of specific microbial taxa in nature.
Publication:
Zhong, K. X., Wirth, J. F., Chan, A. M. & Suttle, C. A. Mortality by ribosomal sequencing (MoRS) provides a window into taxon-specific cell lysis. ISME J (2022). https://doi.org/10.1038/s41396-022-01327-3.
References
- Bergh, O., Børsheim, K. Y., Bratbak, G. & Heldal, M. High abundance of viruses found in aquatic environments. Nature 340, 467-468 (1989).
- Proctor, L. M. & Fuhrman, J. A. Viral mortality of marine bacteria and cyanobacteria. Nature 343, 60-62 (1990).
- Suttle, C. A., Chan, A. M. & Cottrell, M. T. Infection of phytoplankton by viruses and reduction of primary productivity. Nature 347, 467-469 (1990).
- Suttle, C. A. The significance of viruses to mortality in aquatic microbial communities. Microb Ecol 28, 237–243 (1994).
- Suttle, C. A. Viruses in the sea. Nature 437, 356–361 (2005).
- Datta, A. K & Burma, D. P. Association of ribonuclease I with ribosomes and their subunits. J Biol Chem 247, 6795–6801 (1972).
- Deutscher, M. P. Maturation and degradation of ribosomal RNA in bacteria. Prog Mol Biol Transl Sci 85, 369–391 (2009).
- Fuhrman, J. A. & Noble, R. T. Viruses and protists cause similar bacterial mortality in coastal seawater. Limnol Oceanogr 40, 1236–1242 (1995).
- Mojica, K. D. A. & Brussaard, C. P. D. Significance of viral activity for regulating heterotrophic prokaryote community dynamics along a meridional gradient of stratification in the Northeast Atlantic Ocean. Viruses 12, 1293 (2020).
- Bayles, K. W. Bacterial programmed cell death: making sense of a paradox. Nat Rev Microbiol 12, 63–69 (2014).
- Sockett, R. E. Predatory lifestyle of Bdellovibrio bacteriovorus. Annu Rev Microbiol 63, 523–539 (2009).
- Chao, L. & Levin, B. R. Structured habitats and the evolution of anticompetitor toxins in bacteria. Proc Natl Acad Sci USA 78, 6324–6328 (1981).
- Granato, E. T., Meiller-Legrand, T.A. & Foster, K.R. The Evolution and Ecology of Bacterial Warfare. Curr Biol 29, R521–R537 (2019).
- Weitz, J. S. Stock, C. A., Wilhelm, S. W., Bourouiba, L. et al. A multitrophic model to quantify the effects of marine viruses on microbial food webs and ecosystem processes. ISME J 9, 1352-1364 (2015).
- Suttle, C. A. Marine viruses—major players in the global ecosystem. Nat Rev Microbiol 5, 801-812 (2007).
- Thingstad, T. F. Elements of a theory for the mechanisms controlling abundance, diversity, and biogeochemical role of lytic bacterial viruses in aquatic systems. Limnol Oceanogr 45, 1320–1328 (2000).
- Våge, S., Storesund, J. E. & Thingstad, T. F. SAR11 viruses and defensive host strains. Nature 499, E3-4 (2013).
- Bouvier, T. & del Giorgio P. A. Key role of selective viral-induced mortality in determining marine bacterial community composition. Environ Microbiol 9, 287–297 (2007).

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in