First come, first served? Priority effects in infant gut microbiota assembly
Published in Microbiology

The establishment of our gut microbiota in early life plays a significant role in current and future health and disease [1]. However, we still do not fully understand what governs species selection in the gut. Priority effects describe the phenomenon that the timing and order of arrival of species into an ecosystem dictate its composition and function. Indeed, very little is known about the role of priority effects in early life gut microbiota assembly – and how deterministic factors such as diet would influence this.
While both working in the Lab of Professor Tine Rask Licht at the Technical University of Denmark, my colleague, Henrik Munch Roager (now University of Copenhagen), and I established the Danish infant cohort “Copenhagen Infant Gut” back in 2017. This cohort consist of 25 healthy breast-or mixed-fed infants that were fecal sampled longitudinally (9-11 samples per infant) from birth to 6 months of age. We profiled the microbiota in these samples using 16S rRNA gene amplicon sequencing. Our first observation was the striking dominance of Bifidobacterium species in the gut of these infants. Bifidobacterium species are archetypical members of the infant’s gut microbiota owing to their ability to utilize so-called human milk oligosaccharides (HMOs) found in maternal breast milk. These HMO-consuming taxa include B. longum subsp. infantis, B. longum subsp. longum, B. breve, B. bifidum, and B. pseudocatenulatum, of which the former exhibits efficient and broad-range utilization of these oligosaccharides [2].
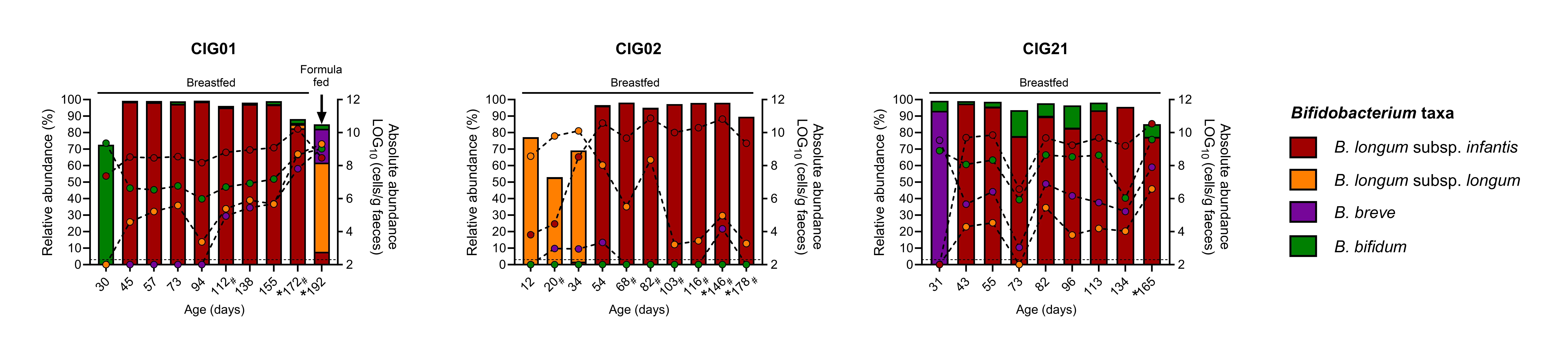
In our cohort, we further investigated the Bifidobacterium species composition using species and subspecies-specific primers and qPCR. Here we discovered that not only were most of these infants packed with HMO-loving species, but individual infants often shifted their dominant Bifidobacterium species/subspecies over time. A particular peculiar phenomenon was observed for B. longum subsp. infantis, which was initially undetected or very low abundant, but became the single dominating taxa in 44% of the cohort infants, somewhere between 14-100 days of life. Intriguingly, these infants were initially colonized and heavily dominated by other Bifidobacterium species, such as B. breve, B. bifidum, and B. longum subsp. longum, as exemplified in three infants in Fig. 1.
.jpg)
This made us wonder if the early arriving Bifidobacterium species were initially benefitting from “seeding first”, but subsequently outcompeted by more superior HMO-utilizers such as B. longum subsp. infantis. Profiling the HMO residuals (originating from the maternal milk) in the infant’s feces gave us a clue. The major HMO structures namely, fucosyllactoses, sialyllactoses, and lacto-N-(neo)tetraoses all exhibited rapid declines, coinciding with the observed temporal increase in B. longum subsp. infantis.
We next designed an animal study to test the hypothesis that priority effects exist in vivo between closely related Bifidobacterium taxa colonizing the gut – and that breast milk-derived HMOs work as selective forces to modify the strength of such priority effects. We sequentially colonized germ-free mice with B. longum subsp. longum, a suboptimal HMO-utilizer, and then B. longum subsp. infantis, as an example of a vivid HMO-utilizer. In a new group of mice, we then reversed the colonization order. We then included two more groups of mice following the exact same schedule, except that they were supplemented with drinking water containing a mixture of HMOs (Fig. 2).
.png)
We observed clear priority effects when mice consumed normal drinking water with the first arriving (sub)species dominating by the end of the study. In contrast, when mice consumed HMO-supplemented drinking water, the superior HMO-utilizer B. longum subsp. infantis always dominated – regardless of the colonization order (Fig. 3). Interestingly, the dominance was more pronounced when B. longum subsp. infantis colonized first!
.jpg)
Our study strongly suggests that priority effects are at play in the early establishment of the infant gut microbiota. Nonetheless, breastfeeding seems to be such a strong deterministic force, driving (sub!) species selection, despite initial priority effects in the infant gut microbiota.
References
- Laursen MF, Bahl MI, Licht TR. Settlers of our inner surface – factors shaping the gut microbiota from birth to toddlerhood. FEMS Microbiol Rev 2021; 001: 1–14.
- Sakanaka M, Gotoh A, Yoshida K, Odamaki T, Koguchi H, Xiao JZ, et al. Varied pathways of infant gut-associated Bifidobacterium to assimilate human milk oligosaccharides: Prevalence of the gene set and its correlation with bifidobacteria-rich microbiota formation. Nutrients 2020; 12: 1–21.



Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in