
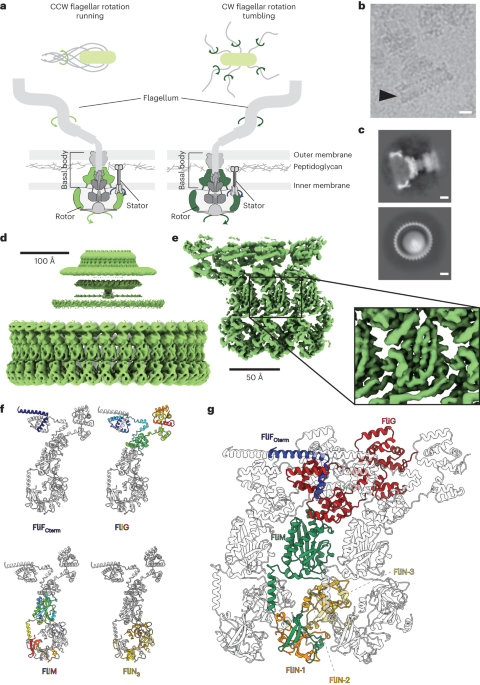
The ability to move towards a food source, or move away from a threat, is a key attribute of successful organisms from across the tree of life. As such, motility has been intensively studied in bacteria for decades, if not centuries; with the bacterial flagellum being at the forefront of this research. In the early 1970s it was recognized by Howard Berg and peers that flagella driven motility was based on the rotation of the flagellar filament. Although there is a large degree of structural variation across different species of bacteria, the core mechanisms seem to be conserved. Filament rotation occurs because it is coupled to a large cytoplasmic ring structure, termed the C-ring, with the rotation of the C-ring being driven via interactions with peripherally associated protein complexes termed stators. Furthermore, chemotaxis, i.e. the ability to change the direction of swimming in response to an attractant or repellant, was shown to be based on the ability of the rotary motor to switch the direction of rotation. Massive strides have been taken in the last few years when it comes to understanding the detailed architecture of much of the flagellar basal body, largely thanks to the opportunity to study large, intact complexes with cryo-electron microscopy (cryoEM). The recent revelation that the stator complexes are themselves rotary motors led to a new model of flagellar rotation, in which the large C-ring “cog” is a passive motor component, being actively driven by one or more smaller stator cogs at the periphery. But this model begat new questions about the mechanisms by which binding of chemotactic regulators to the passive cog could alter the direction of C-ring rotation, despite no alteration to the direction of rotation of the active stator cogs; with one proposal being that structural changes in the C-ring could lead to a change in the passive cog “teeth” directionality, and subsequent stator relocation (Figure 1).
Our lab’s interest in motility originally stemmed from a more general interest in type III secretion systems, one example of which lies at the heart of the bacterial flagellar basal body. After solving structures of some the inner-membrane tethered complexes via heterologous expression (described in https://communities.springernature.com/posts/one-ring-to-rule-them-all), we decided in early 2020 to have a go at extracting intact basal bodies directly from Salmonella cells, partly inspired by the observation that the stoichiometry of these highly symmetric structures could be influenced by the presence or absence of interacting components. And……. then the pandemic struck. After a couple of months of almost complete lockdown, staff were cautiously allowed back into the lab, under strict distancing guidelines that required a complex dance of experimental planning. Against all the odds we were able to collect good micrographs in the summer and early autumn of 2020, leading to structures of the cell envelope spanning portions of the complete basal body (https://www.nature.com/articles/s41564-021-00895-y). These structures provided great insight into the assembly of this immensely complex object, including how many different symmetries interweaved to form the final structure. However, the portions of the flagellum responsible for actual flagellar rotation, i.e. the C-ring and the peripheral stators, were missing from the final reconstructions, and so we could say little about the core function of this molecular machine at this point.
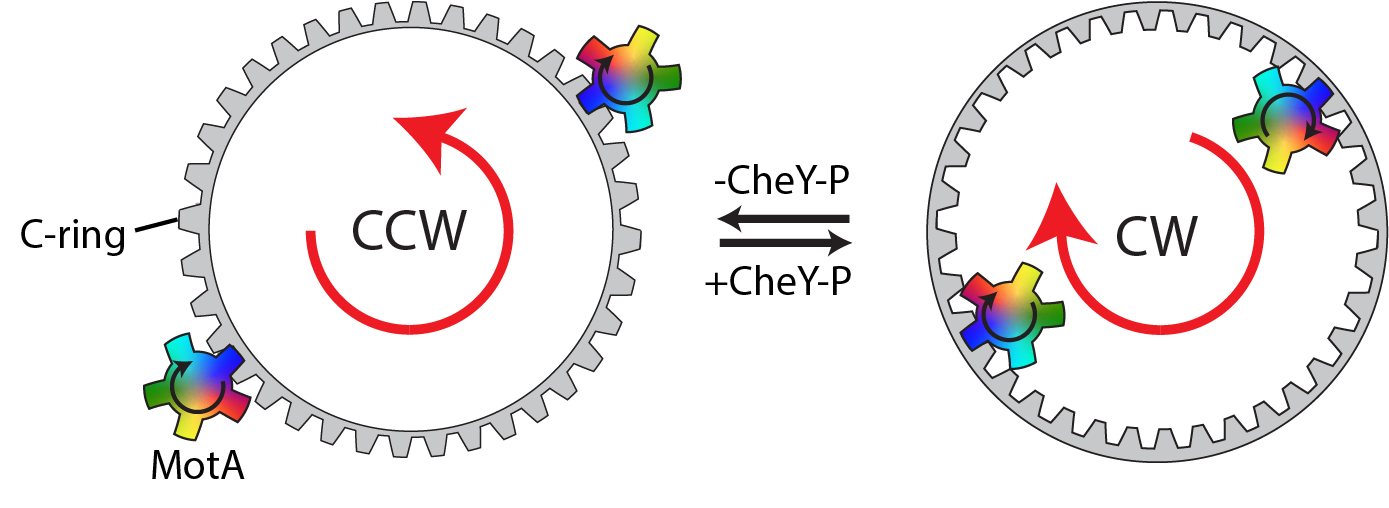
Just to make 2020 even more complex, the lab was also in the process of moving continents, from the dreaming spires of Oxford (UK) to the clustered spires of Frederick (USA), home of a campus of the National Cancer Institute. Of course this was still mid-pandemic, so getting visas and making travel arrangements was challenging, but by Christmas 2020 several lab members were safely ensconced in their new home in the US. And thus began the rather complex task of building a lab from scratch, in a new country, with physical access to the lab restricted, and backlogs on equipment ordering due to pandemic supply chain issues. While the extended lab set-up period was frustrating, it did provide us with the time to go back and re-analyze old datasets with updated versions of the software packages. Out of that analysis we were able to identify a subset of basal body particles that retained intact C-rings. Furthermore, we had collected data from mutants trapped in counterclockwise (CCW) and clockwise (CW) rotation modes, giving us the capacity to compare the structural details of the two states. These analyses revealed extensive global changes in the C-ring, but crucially demonstrated 180 degree rotations of two distinct structural domains of the C-ring protein (FliG) that interacts with the stators. However, to fully understand the implications of this for the mechanism of rotation reversal, we needed a clearer picture of the structural basis of the stator interactions with the C-ring. This was achieved by fusing the stator interaction domain of FliG to the C-terminus of the major stator component (MotA), thus stabilizing the relatively weak interaction between stator and C-ring, and producing a high resolution structure of the important interaction. By docking this structure onto our two C-ring conformations we were able to build up snapshots of the CCW and CW C-rings in their stator engaged states, demonstrating that remodeling of the FliG protein alters the teeth of the C-ring and hence the stator interaction surface, thus validating our earlier model. In the default pose, stators interact with the external face, and CW rotation of the small cog drives CCW rotation of the larger cog. Binding of the chemotactic regulator to the C-ring alters the structure such that the stator now interacts with the inner face, and thus the same CW small cog rotation now rotates the large cog in a CW direction (Movie 1).
Movie 1: Animation demonstrating gearing in the Salmonella flagellum. The small stator cog (rainbow) rotates in response to the proton-motive force (pmf). The teeth of the small cog drive rotation of the large C-ring cog (gray), which in turn drives the rotation of the flagellum. Structural transitions of the C-ring in response to chemotaxis regulators remodel the teeth of the large cog, reversing the direction of rotation.
The structures also provided insight into the high degree of cooperativity of the regulator induced switching. Firstly, the rotation of the stator pushes the C-ring subunits in the direction of their switched conformation, meaning they are primed for the switch event. Secondly, the C-ring subunits are intricately inter-twined, and lateral shifts sterically preclude neighboring subunits existing in different states, therefore regulator induced changes propagate rapidly around the ring from a single binding site. These elegant mechanisms provide exquisite sensitivity to environmental signals, and also minimize the chance of having opposing forces operating on different sides of the C-ring cog, i.e. rotational switching is “all or nothing”. It seems like the more we learn about this remarkable molecular machine, the more awe inspiring it becomes.
Follow the Topic
-
Nature Microbiology
An online-only monthly journal interested in all aspects of microorganisms, be it their evolution, physiology and cell biology; their interactions with each other, with a host or with an environment; or their societal significance.


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in