Raiders of the Lost Rudiment: ancient ‘useless’ structure makes ants huge
Published in Ecology & Evolution


In 2002, a now classic paper was published that used ants as a model system to understand the dynamics and evolution of gene regulatory networks in the context of developmental plasticity (Abouheif & Wray, 2002). While the reproductive castes in ant colonies (queens and males) have wings for mating and migration, all known extinct and extant worker ants lack wings. In holometabolous insects, which include ants, wings develop from populations of cells called imaginal discs. Despite this universal absence, there are two very peculiar things about developing ant workers: (1) they actually transiently develop wing imaginal discs; (2) the degree to which this tissue grows and patterns in a developing worker differs within and between species.
At the end of Abouheif & Wray (2002) the question was asked: is the variation of these rudiments neutral and the result of drift (as the adults lack wings), or is there something more to their existence; perhaps this variation is somehow the result of selection?
From the start of my grad studies back in the mid-late 2000's many observations fuelled this burning question. At first we worked within a framework that assumed the differences in rudimentary wing discs was truly just a byproduct of differences in hormonal levels between developing winged castes and wingless castes. Inspired by work in Drosophila that spanned from Beadle & Ephrussi in the 1930's, to Schubiger, Bryant, Simpson, as well as many others in the 1980's and 90's, we began to shift towards the hypothesis that perhaps the rudimentary wing imaginal discs may in fact do something when they grow.
This was further solidified by preliminary phylogenetic studies that we were conducting, which demonstrated that there was an association between the degree of rudimentary wing disc development and the occurrence of an soldier caste. Finally, we observed that the rudimentary wing disc of Pheidole soldier larvae proliferate quite extensively and then surprisingly undergo finely-patterned apoptosis, and degrades prior to metamorphosis. All that being said, one thing for sure, this all certainly culminated into a eureka moment, where we are sitting there, in Ehab Abouheif's office exclaiming: "oh my god, what if they have a function?!" This is the prime question that we aimed to address in our paper published in Nature today.
We first set out by targeting the soldier rudimentary wing disc in the species Pheidole hyatti using larval RNAi, which we had to develop for the ants. Long before gearing up for this, we were not too keen about taking that approach, and then Andrew Fire came and visited our lab during a seminar day at McGill back in 2007, when we talked about the dream of doing RNAi in ants...inspiring would be one of the words I would use. We targeted a gene named vestigial, which has been extensively studied in Drosophila and is known to be the master identity regulator for wings, and both necessary and sufficient for wing development. Based on the fly work from the 1980's we thought, maybe it might change the size of the individual.
The RNAi was a success in terms of specifically perturbing the development of the wing disc, giving a range of phenotypes from completely suppressed growth to mildly perturbed. So, after the RNAi injections, we we planned on measuring individuals to see if they were bigger or smaller. A fellow graduate student (Marie-Julie Fave) and I took some individuals out of the experimental replicates and placed them under the microscope. WHAT?! Oh my...they looked like little microsoldiers?! We began to look at more and we started to realize that we created a whole series of individuals with varying degrees of head proportions and body size, where we found that a more reduced body size was accompanied by an even greater reduction in head size.
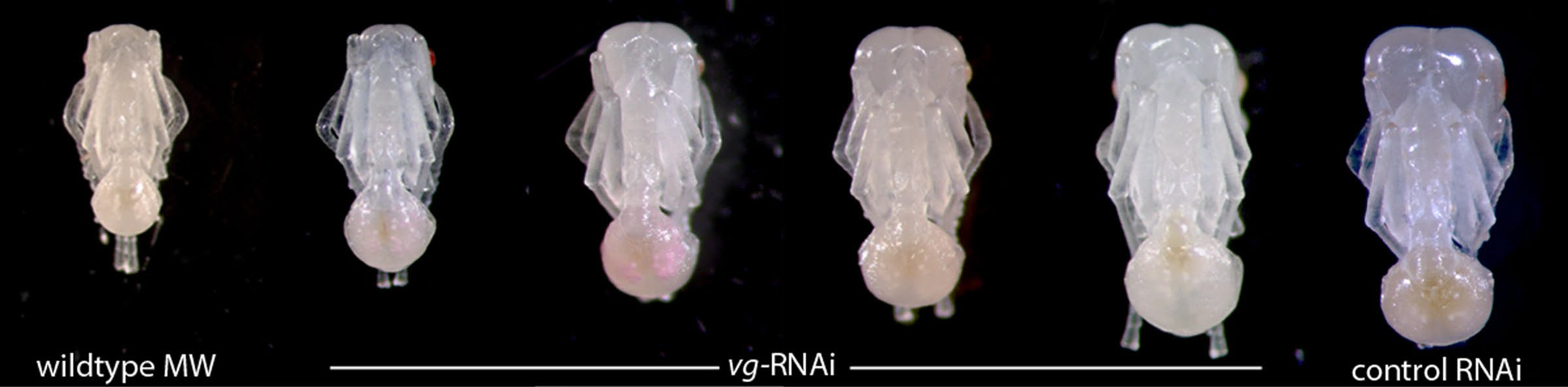
MJ and I freaked out, practically doing cartwheels in the hallway. I mean, we knew, almost immediately, that we found something worth doing cartwheels for. How often do you see something so unexpected yet immediately feel equally gratified. This is science. Usually, that moment is followed by shear fear, like, no no no, this cannot be right. But, in all honesty, we went into the experiment with a, who-knows-what-we'll-find attitude...and it was what it was.
It stems from our mentor, Ehab Abouheif, who instilled in us a kind of Indiana Jones-style science: blaze through the literature in seek of insight, tinker in the lab and manipulate the experimental systems and, most of all, go out into the field with Curiosity in one hand and Crazy in the other, and let nature provide the questions for you. With this cocktail, serendipity will follow.

One thing to add to that point, even Indiana Jones had inspiring figures to look up to, and over the years the Abouheif lab, myself included, have benefitted an immense deal from the wealth of ant knowledge that various myrmecologists (and true natural historians) have provided us, including Ray Sanwald, Robert Johnson, Lloyd Davis, and Walter Tschinkel.

Back to the paper. We later confirmed these RNAi results with other techniques, including ablations, where we (Sophie Koch) zapped the developing wing imaginal discs. Furthermore, we demonstrated that the function of the developing wing disc was specific to the soldier caste. At this point, it was pretty clear to us that rudimentary wing discs were important in regulating head-to-body allometry. Still, this finding literally opened the door to an even crazier idea:
In the 1980s, Wheeler & Nijhout discovered that in the genus Pheidole, the insect hormone juvenile hormone (JH) is the critical growth hormone that can induce soldiers when at high levels or minor workers when low. Subsequently, they did social manipulation experiments that demonstrated that, in addition to this positive soldier-caste determining factor, larvae destined to become soldiers can be inhibited if raised in an environment that was highly populated by adult soldiers. With detailed experiments and observations, they proposed that this was caused by an inhibitory pheromone, which blocked soldier development. To us it was obvious: raise soldier-destined larvae with adult soldiers and see if that inhibitory pheromonal environment would affect the disc. It turns out the pheromone recapitulates the results we obtained from the RNAi experiments: a range of allometrically-sized individuals caused by a range of suppression of rudimentary wing disc development.
We then isolated the soldier inhibitory pheromone and repeated the experiments, demonstrating that it was not that soldiers were bad caregivers, rather it was indeed the pheromone at play. Finally, we tried a fantasy experiment: what happens when an unstoppable force (JH) meets an immovable object (soldier inhibitory pheromone). We could not have predicted the outcome: we generated the very beginnings of a soldier-destined individual, they were barely allometric and they developed from larvae that just started to grow rudimentary wing discs but did not cross the caste determination threshold. Based on all of our pheromone and juvenile hormone experiments, we came to the conclusion that the pheromone primarily acts by suppressing soldier rudimentary wing disc development and that juvenile hormone both induces the allometric aspects of soldier development by activating rudimentary wing disc development and can increase overall body size in a proportional way independent of the disc.
Incredibly, when you take our experimental data together, do an allometry plot, and compare it to the natural Pheidole hyatti plot, you see something completely unexpected: by perturbing the rudimentary wing disc’s development, we generated lost phenotypic variation that recapitulates the allometry found in other distantly related ant species. Furthermore, the soldier larvae of other species that have independently evolved a soldier caste, have massive rudimentary wing discs. This brings me to what we now call 'Wilson's transitions'.

Darwin was baffled by the morphological diversity among the various worker subcastes that he observed within an ant colony, and to paraphrase, he thought it was almost fatal to his theory of natural selection. Among other things, how could you have big-headed soldiers that use their specialized morphology to defend the colony and completely different small-headed workers optimized for foraging and nursing young. Of course he had an incredible hypothesis to explain it, this later inspired a masterwork by E.O. Wilson.
Fast forward to 1953, E.O. Wilson, based on Darwin's observation, and further inspired by the writings and analysis of growth and allometry by Julian Huxley in 'Relative Problems of Growth' (1932), attempted to propose a framework to understand how a complex worker-caste system can evolve, which could have 2 or even 3 worker subcastes, from an ancestral species that started where all worker ants looked the same (a must read). Wilson used Huxley's allometry plots to map out the distribution of individual traits, especially head size, in relation to body size across whole colonies of ants for several species. Based on his data he proposed a framework by which you can take a distribution of workers that is initially linear, introduce non-linear properties to it, followed by selection on extreme phenotypes that would lead to bimodal and perhaps even trimodal-like distributions of worker castes that are specialized for particular tasks.
This is why we have proposed that the rudimentary wing discs, in developing soldiers across ants (and perhaps to a more subtle degree in other worker caste-systems) is the key mediator that facilitates what we call: 'Wilson's transitions'. Selection on the degree of development and formation of these rudimentary structures can increase (or decrease) the allometric variation amongst worker individuals. Alongside JH, and likely several other growth factors and signalling molecules, the rudimentary discs may facilitate the transitions of the worker caste from a simple monomorphic system, to a more complex dimorphic one.

This work further illuminated our previous work that demonstrated that JH can re-activate dormant developmental potentials to produce ancient supersoldier phenotypes (Rajakumar et al., 2012). In Pheidole, supersoldiers, which are even more allometrically disproportionate than soldiers, have really big rudimentary wing discs (including an additional 2nd pair) that are activated by an additional JH threshold late in larval development. Our new work now provides the mechanism by which we were able to induced supersoldiers in species that lack them: we activated the development of a 2nd pair of wing discs, which triggered the more elaborate allometric growth akin to supersoldiers. Again, until now, we had thought this 2nd pair of wing discs was just a defining developmental marker for the caste, now we know it is the defining regulatory tissue.
More generally, we propose that rudiments may provide an additional source for natural selection to act upon to generate parallel evolution and novelty. In the case of ants, having a worker caste that develop wing imaginal discs, which no longer are constrained to produce a functional wing, enables wing rudiments to elaborate their more ancient interorgan communication systems such that they can directly or indirectly cause the head to grow disproportionally larger. Subsequently, in Pheidole, these rudiments become sensitive to the social environment and novel inhibitory pheromones.
Who knows what additional unexpected regulatory roles rudiments play in other organisms (including us), it is certainly worth thinking about.




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in