Regulation of VHL-mediated HIF-1α protein degradation under normoxia —a potential target in cancer treatment
Published in Cancer
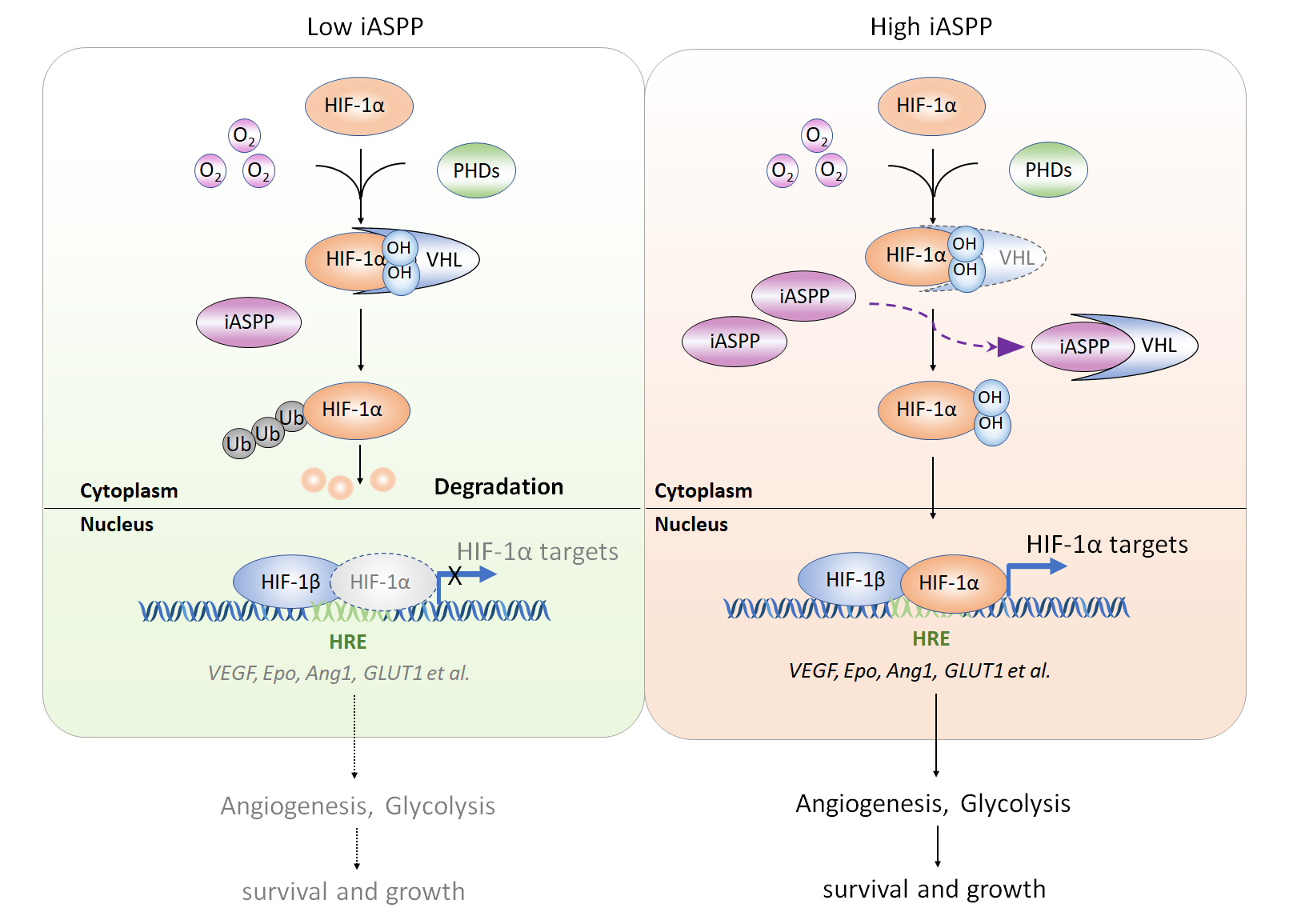
Hypoxia-inducible factor-1α (HIF-1α) is a master regulator of hypoxic adaption. It is frequently overexpressed in human cancers; however, its expression is not always associated with hypoxia in vivo. Although loss of von Hippel–Lindau (VHL) is the best known mechanism that ensures constitutive activation of HIF-1α in clear cell renal cell carcinoma, little is known about how VHL wild-type tumors acquire the ability to constitutively activate HIF-1α. This is a critical question, because HIF-1α activation under normoxia leads to a pseudohypoxic response, which manifests as altered transcriptional regulation of numerous HIF targets leading to carcinogenesis. Therefore, better understanding of the oxygen-independent constitutive activation of HIF-1α could shed new light on cancer pathology and disease mechanisms, and may facilitate the development of new anticancer therapies.
iASPP (inhibitor of apoptosis-stimulating protein of p53) is the most conserved member of the ASPP family[1]. It has been generally assumed that the oncogenic function of iASPP was fulfilled mainly through directly binding with p53 (or its family members p63 and p73) and selectively inhibiting their transcriptional activities on their proapoptotic targets[2, 3]. Whereas our recent studies demonstrate that the oncogenic effects of iASPP can be p53-independent. For instance, iASPP inhibits apoptosis by activating the antioxidative transcription factor Nrf2 and confers drug resistance in renal cell carcinoma[4]. iASPP overexpression confers resistance to cytosolic Ca2+-mediated apoptosis in colon cancers by stabilizing Ca2+ channel protein TMCO1[5]. It is likely that iASPP is an essential anti-apoptotic regulator involved multiple mechanisms. Also notably, iASPP is overexpressed in multiple types of cancer, and increased iASPP expression is associated with diverse malignant phenotypes such as tumor growth and metastasis. Resistance to apoptosis may not be able to explain all iASPP-linked oncogenic phenotypes, and whether iASPP is involved in the regulating of other cancer hallmarks is largely unknown. With this question in mind, we launched a study, which leads to the discovery of a novel mechanism through which HIF-1α-mediated glycolysis and angiogenesis is activated by iASPP overexpression under normoxia.
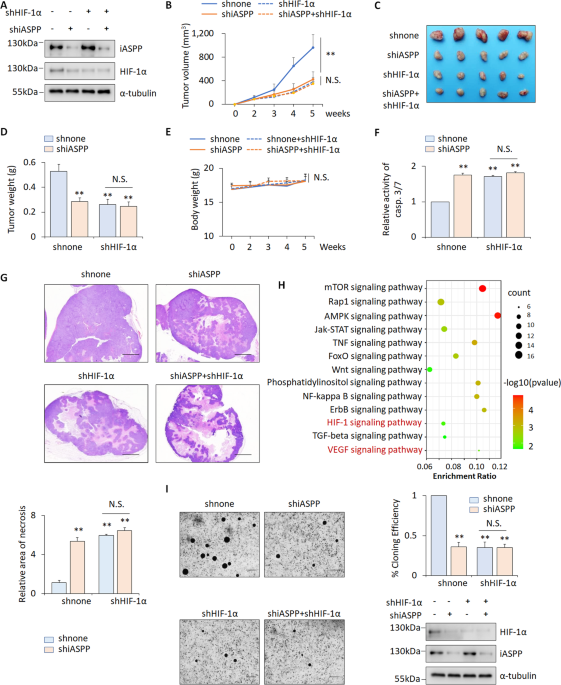
Initially, we noted that, despite the smaller size of iASPP knockdown (KD) xenografts, the proportions that were necrotic were significantly higher in iASPP KD xenografts compared with controls, suggesting that the inhibitory effect of iASPP on tumor growth in vivo may be at least partially due to intolerance to hypoxia. This was subsequently validated in HIF-1α KD xenografts, demonstrating that the pro-tumor effect of iASPP can be abolished by HIF-1α KD. In addition, bioinformatic analysis of the RNA-seq data derived from the control and iASPP-overexpressing cells reveal that iASPP is indeed associated with HIF signaling. Further functional studies show that iASPP promotes angiogenesis and glycolysis in a HIF-1α dependent manner.
If iASPP is essential in hypoxic adaption, its expression levels may be changed with oxygen concentration. Nonetheless, it is not the case. The expression levels of iASPP are comparable under normoxia and hypoxia. Also unexpectedly, iASPP regulates angiogenesis and glycolysis under both normoxia and hypoxia. Similarly, iASPP promotes hypoxic-response element (HRE) luciferase activity and HIF-1α targets gene expression regardless of oxygen concentration in a HIF-1α-dependent manner. These observations motivate us to explore whether iASPP is an oncogene that confers constructive activation of HIF-1α regardless of oxygen concentration.
It should be noted that HIF-1α is unstable with a half-life about 5 minutes under normoxia in most cell lines tested. The protein levels of HIF-1α are barely detected under normoxia, which made it difficult to evaluate the impacts of iASPP on HIF-1α expression under normoxia. We then modified the conventional western blot assay by increasing the protein loading amount to 80μg. Under such conditions, we were able to consistently detect HIF-1α expression in nomoxic cells. In addition, we found that iASPP overexpression results in increased HIF-1α protein, while iASPP KD decreased it under both normoxia and hypoxia (2% oxygen). In contrast, HIF-1α mRNA levels are not changed with iASPP overexpression or KD.
We then set out to investigate how iASPP activates HIF-1α by increasing its protein levels. It turns out that iASPP acts mainly by inhibiting proteasome-mediated HIF-1α degradation by interfering with the PHD/VHL pathway. Mechanistically, (1-294) iASPP contains key VHL-binding residues and iASPP bind with VHL mainly via residues mapped to 1-154 aa in VHL. (1-154) VHL contains a key HIF-1α-binding residue, Y98[6]. Interestingly, the ability of Y98N-VHL to bind with iASPP is also dramatically reduced, suggesting that Y98N in VHL is a key residue in its binding with both HIF-1α and iASPP. Our data show further that binding of VHL with iASPP prevents it from binding with HIF-1α, leading to increased stability of HIF-1α protein.
Above findings are mainly derived from cell culture systems. To understand the roles of iASPP-HIF-1α axis in vivo, we further examined the expression levels of iASPP, HIF-1α and the key downstream targets of HIF-1α in colon cancer tissues and their compared normal controls. The positive association between iASPP, HIF-1α protein and VEGF and GLUT1 were observed, which further suggest that iASPP-HIF-1α axis may occurs in vivo. Indeed, iASPP-regulated HIF-1α stabilization was validated in xenograft models. Moreover, the animal model also revealed that iASPP KD xenografts were more sensitive to YC-1 (HIF-1α inhibitor) than the shnone controls, indicating that iASPP inhibition can enhance the chemotherapeutic efficacy of YC-1 (low toxic dose) by further suppressing HIF-1α expression in cancer cells.
This study is important, because it establishes a novel mechanism that regulates HIF-1α’s constitutive activation in cancers under normoxia. In addition, it extends iASPP’s functions from apoptosis regulation to angiogenesis and glycolysis. Furthermore, inhibition of the iASPP-HIF-1α axis inhibits angiogenesis and glycolysis both in vitro and in vivo, highlighting the potential of the iASPP-HIF-1α axis as an important anticancer treatment target.
References
1. Bergamaschi, D., et al., iASPP oncoprotein is a key inhibitor of p53 conserved from worm to human. Nat Genet, 2003. 33(2): p. 162-7.
2. Lu, M., et al., Restoring p53 function in human melanoma cells by inhibiting MDM2 and cyclin B1/CDK1-phosphorylated nuclear iASPP. Cancer Cell, 2013. 23(5): p. 618-33.
3. Cai, Y., et al., iASPP inhibits p53-independent apoptosis by inhibiting transcriptional activity of p63/p73 on promoters of proapoptotic genes. Apoptosis, 2012. 17(8): p. 777-83.
4. Ge, W., et al., iASPP Is an Antioxidative Factor and Drives Cancer Growth and Drug Resistance by Competing with Nrf2 for Keap1 Binding. Cancer Cell, 2017. 32(5): p. 561-573.e6.
5. Zheng, S. and D. Zhao, iASPP suppresses Gp78-mediated TMCO1 degradation to maintain Ca(2+) homeostasis and control tumor growth and drug resistance. Proc Natl Acad Sci U S A. 2022. 119(6).
6. Tanimoto, K., et al., Mechanism of regulation of the hypoxia-inducible factor-1 alpha by the von Hippel-Lindau tumor suppressor protein. Embo j, 2000. 19(16): p. 4298-309.
Follow the Topic
-
Oncogene
This journal aims to make substantial advances in our knowledge of processes that contribute to cancer by publishing outstanding research.


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in