Serendipity in the balance: an unexpected finding in PHA production by purple phototrophic bacteria
Published in Bioengineering & Biotechnology, Chemistry, and Sustainability
Some discoveries start with a grand hypothesis. This one started with an annoying mismatch.
We were not looking for crotonate. We were trying to optimize the production of PHA, a biodegradable polymer that bacteria can store inside their cells. In simple terms, PHA is one of the microbial routes that could help us move away from fossil-based plastics. Purple phototrophic bacteria are attractive for this because they can use light as an energy source and convert organic carbon into valuable products with remarkable efficiency.
So the original question was straightforward: can we push these bacteria to accumulate more PHA?
As chemical engineers, we approached the system in the way we know best. We measured substrates, biomass, nitrogen, polymers, hydrogen, gases, and metabolites. We tried to close the mass and electron balances. Carbon should go somewhere. Electrons should go somewhere. If acetate and butyrate disappear from the medium, they should end up as biomass, PHA, CO₂, hydrogen, or another measurable product.
But there was a loose end.
At first, it looked like analytical noise. A small signal appeared in the supernatant, not where we expected the usual compounds. It was not the main product. It was not even a large fraction of the carbon. But it was there. And, more importantly, it appeared exactly when the cells were under metabolic pressure.
The molecule was crotonate.
That was surprising. Crotonate is not something we expected wild-type purple phototrophic bacteria to release into the medium. It is chemically close to central intermediates involved in PHA metabolism, but extracellular crotonate release had mostly been associated with engineered organisms. In our case, the bacteria had not been designed to do this. They were just reacting to the conditions we imposed.
At that point, the story became less about maximizing PHA and more about understanding why the bacteria were “leaking” this molecule.
We first had to be sure we were not fooling ourselves. The most dangerous discoveries are the ones that fit a nice story too early. So we did the boring but essential work: standards, retention times, fragmentation patterns, repeated checks by GC MS and GC MS/MS. We asked whether the signal could come from contamination, misidentification, sample handling, or a hidden compound behaving similarly. Only after several sanity checks did we accept that the molecule in the supernatant was indeed crotonate.
Then came the real question: why would the bacteria release it?
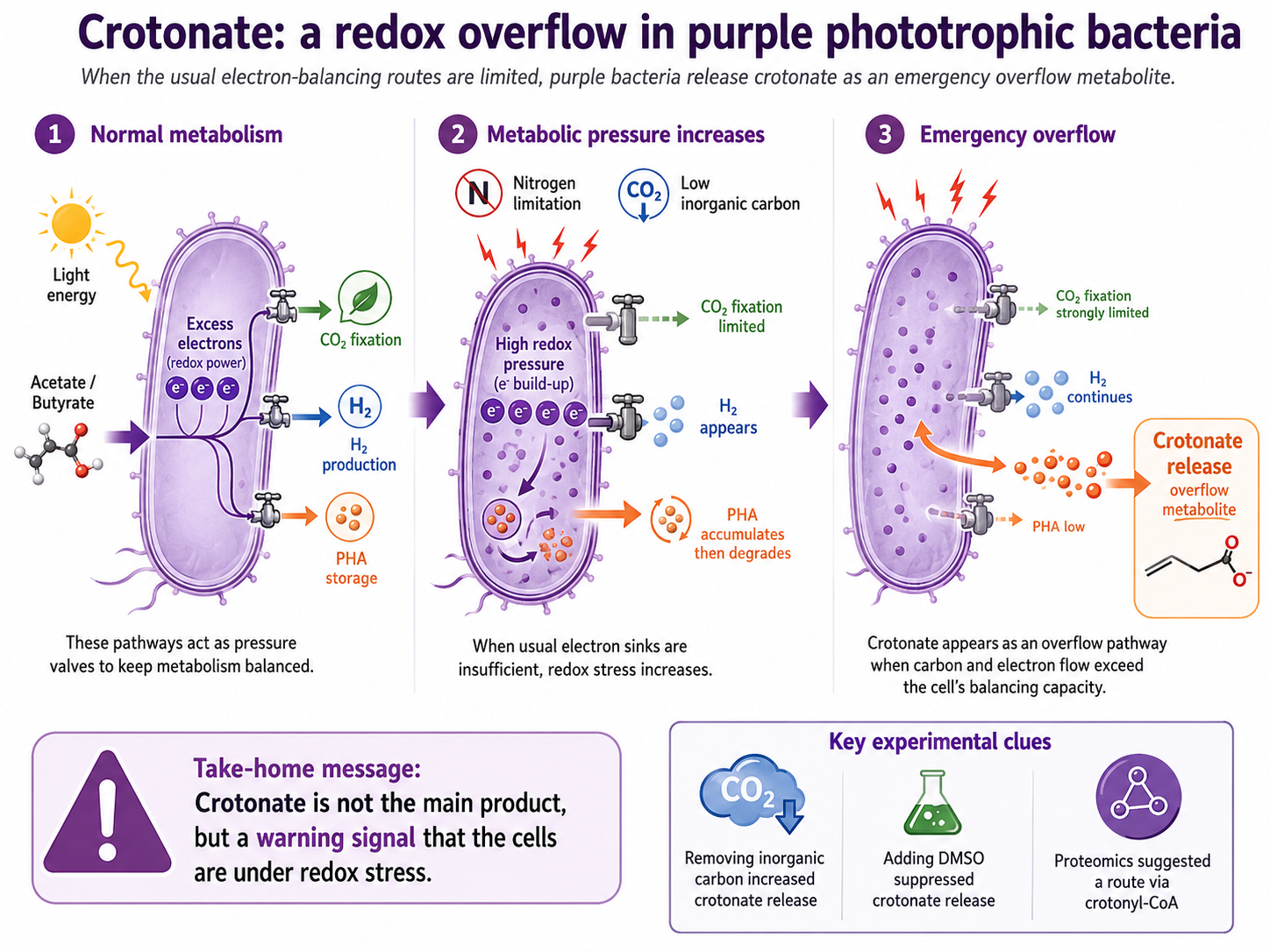
Purple phototrophic bacteria are experts in redox balancing. When they grow on reduced organic substrates under light, they must constantly manage excess electrons. They have several ways to do this. They can fix CO₂. They can produce hydrogen. They can store carbon and electrons as PHA. These routes are like pressure valves that keep metabolism running.
But what happens when these valves are limited?
Our experiments suggested that crotonate appears when the usual routes are not enough. Under nitrogen limitation, growth slows down and the cells cannot keep converting carbon into new biomass. PHA first accumulates, but later it is degraded. Hydrogen production appears as another outlet. And then crotonate shows up in the liquid phase.
To test whether this was really linked to redox stress, we made the “headache” worse for the bacteria. We removed inorganic carbon, which limits one of their major electron sinks: CO₂ fixation. This is like telling the cells: you have too many electrons, but one of your best ways to spend them is blocked. Under these harsher conditions, crotonate release increased strongly.
The analogy we used in the lab was simple: we were giving the bacteria a bigger and bigger metabolic headache. At some point, when they could no longer relieve the pressure through the usual routes, they released crotonate. Not as a main product, not as an efficient production strategy, but as an emergency overflow.
The opposite experiment was just as important. We added DMSO, an alternative electron acceptor. If crotonate was really part of a redox-balancing response, then giving the cells another way to dump electrons should reduce its release. That is what happened. Crotonate disappeared from the medium. This was one of the most convincing moments in the project, because it showed that crotonate was not simply a dead-end waste product. It behaved like a reversible overflow metabolite.
The proteomics gave us a glimpse of what was happening inside the cells. When crotonate release was highest, Rhodopseudomonas bacteria seemed to be especially active. The cells appeared to slow down one of their normal routes for using carbon to build new material, while increasing proteins linked to the formation of crotonyl-CoA, a molecule closely related to PHA metabolism. We also found a possible enzyme (a candidate CoA-transferase) that could transform this intracellular molecule into free crotonate, allowing it to leave the cell. This helped us move from a simple observation (crotonate appearing in the liquid) to a biological explanation: the cells were likely redirecting their metabolism to cope with excess reducing power.
This does not mean we have closed every mechanistic detail. We have not. The next steps should include isotope tracing, pure culture validation, and targeted genetic work to confirm the precise route. But the picture is now much clearer: crotonate release appears to be a previously overlooked overflow pathway in purple phototrophic bacteria, triggered when carbon and electron flow exceed the capacity of the usual balancing mechanisms.
For biotechnology, this matters. If we want to use purple bacteria to produce PHA, protein, pigments, or other products, we need to understand not only what we want them to make, but also what they make when they are stressed. Crotonate may become a useful warning signal. A small molecule telling us that the cells are struggling to keep their metabolism balanced.
In the end, this paper is a reminder that good engineering is not only about optimization. Sometimes it is about accounting carefully enough to notice that something does not add up. And sometimes, behind that missing term in the balance, there is a new piece of biology waiting to be found.
https://www.nature.com/articles/s42003-026-10313-w

Follow the Topic
-
Communications Biology
An open access journal from Nature Portfolio publishing high-quality research, reviews and commentary in all areas of the biological sciences, representing significant advances and bringing new biological insight to a specialized area of research.

What are SDG Topics?
An introduction to Sustainable Development Goals (SDGs) Topics and their role in highlighting sustainable development research.
Continue reading announcementRelated Collections
With Collections, you can get published faster and increase your visibility.
Artificial Intelligence Methodology in Structural Biology
Publishing Model: Hybrid
Deadline: Nov 30, 2026
Healthy Aging
Publishing Model: Open Access
Deadline: Dec 31, 2026

Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in