Uncovering a Metabolic Checkpoint in Autoimmune Inflammation: How Lactate Shapes Immune Hotspots
Published in Biomedical Research
When we think of inflammation in autoimmune diseases, we often picture a generalised immune overreaction. But under the microscope, something far more organised often appears: structured clusters of immune cells known as ectopic lymphoid structures (ELS), which resemble lymph nodes. These formations are a common feature in several chronic conditions, from cancer and cardiovascular disease to autoimmune disorders like rheumatoid arthritis and Sjögren’s disease (SjD). But what governs their formation, persistence, and resolution has remained largely elusive.
Our journey to answering this question began with a surprising suspect: lactate.
The mystery of the inflamed salivary gland
Sjögren’s disease, a systemic autoimmune condition that primarily affects the salivary and tear glands, offered us a uniquely accessible window into ELS formation. Patients often undergo small lip biopsies to aid diagnosis, and these samples revealed striking immune cell clustering, hallmarks of ELS. We were particularly intrigued by the role that CD4+ T cells, a type of immune cell central to inflammation and autoimmunity, played in forming and sustaining these structures.
Building on our previous work showing that lactate influences CD4+ T cell function in rheumatoid arthritis, we asked: Could lactate also be orchestrating the organisation of these immune cell clusters in SjD?
Rethinking lactate: from by-product to bio-signal
Lactate is traditionally thought of as a waste product of metabolism, what muscles produce during exercise or what builds up during hypoxia. But recent research, including our own, suggests that lactate is far from passive. It actively shapes immune cell behaviour: dictating movement, cytokine production, and even cellular metabolism. In chronic inflammation, lactate isn’t just present, it’s abundant.
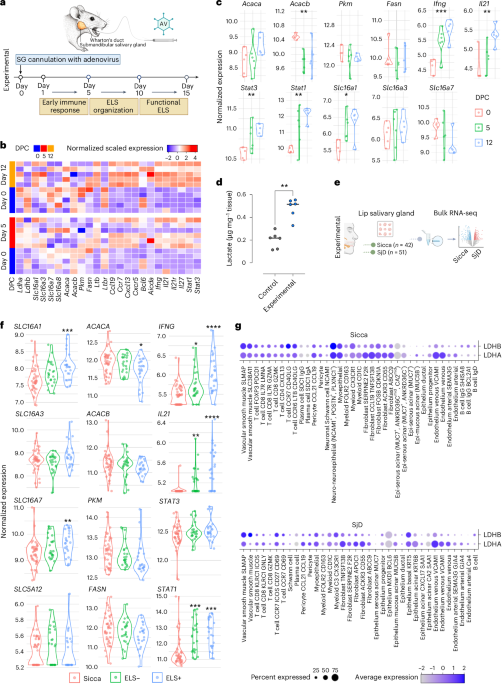
We found that in inflamed salivary glands, lactate concentrations could rise to 10-15 mM, far above the normal range of 1-2 mM. To investigate further, we used a combination of mouse models, human tissue biopsies, single-cell RNA sequencing, and functional assays. What we discovered was a tight link between lactate levels, immune cell architecture, and inflammation.
The transporter that tipped the balance
At the heart of this story lies SLC5A12, a sodium-dependent lactate transporter. Unlike other transporters like MCT1 or MCT4, which are broadly expressed and carry lactate with protons, SLC5A12 is more selective. Under normal conditions, it’s mostly found in the kidneys. But in inflamed tissue, particularly in CD4+ T cells within ELS, it’s dramatically upregulated.
We observed that when SLC5A12 was blocked, either genetically or with a monoclonal antibody (3C7) we developed [IP: University of Birmingham], ELS became less organised, inflammatory cell clusters shrank, and key cytokines like IL-21, crucial for B-cell help, were downregulated. These results weren’t just visible under the microscope; they translated into measurable resolution of inflammation.
Following the signal
In one of the more exciting parts of our study, we tracked how lactate impacted the internal machinery of T cells. Using protein acetylation arrays, we found that lactate exposure changed the expression and modification of hundreds of proteins. One particularly important pathway that emerged was the STAT-IL21 axis, a signalling cascade involved in maintaining germinal centres within ELS.
This was a lightbulb moment. It helped explain why IL-21, a cytokine central to T-B cell cooperation and autoimmunity, was elevated in SjD and how its production was metabolically controlled by lactate uptake through SLC5A12.

From bench to bedside: potential therapeutic implications
One of the major challenges in autoimmune disease treatment is precision, how to calm harmful inflammation without weakening the body’s overall immune defences. Many current therapies, such as systemic immunosuppressants or biologics targeting TNF or B cells, offer broad relief but come with a significant risk of side effects, including increased susceptibility to infections.
What excites us about targeting SLC5A12 is its tissue-specific expression. Unlike many immune receptors or metabolic pathways that are active throughout the body, SLC5A12 is virtually silent in healthy peripheral tissues. Its expression is sharply upregulated only in inflamed tissues, and even more selectively in specific immune cell subsets, like CD4+ T cells and B cells, within those inflammatory hotspots. This makes it an ideal precision target, allowing us to modulate local immune activity while sparing systemic immunity.
Our findings open the possibility that SLC5A12 blockade could serve as a novel metabolic checkpoint therapy in autoimmunity, analogous, in some ways, to checkpoint inhibitors in cancer. But unlike PD-1 or CTLA-4 blockade, which boosts immune responses, targeting SLC5A12 may recalibrate or resolve dysregulated immune circuits that sustain chronic inflammation.
Beyond Sjögren’s disease, this strategy could be extended to other conditions where ectopic lymphoid structures play a role, such as rheumatoid arthritis, multiple sclerosis, systemic lupus erythematosus, and even chronic transplant rejection. In each case, metabolically active immune cells within inflamed tissues may rely on lactate signalling to persist and organise. Blocking that signal could disrupt pathological immune niches without broadly suppressing host defence.
There’s also growing interest in targeting tissue metabolism as a therapeutic axis in its own right. Just as we now recognise the gut microbiome or stromal microenvironment as critical regulators of disease, we’re beginning to appreciate that metabolic context matters. Modulating transporters like SLC5A12 may become part of a new class of therapies, “metabo-immunomodulators”, designed to shift the local biochemical conditions that dictate immune cell fate and function.
Of course, questions remain: How durable are the effects of SLC5A12 blockade? Could it be used in combination with current biologics or immune-modulating drugs? What other cell types might be affected in different disease contexts? These are avenues we’re actively exploring.
The untold stories behind the science
Science, of course, doesn’t happen in a vacuum. There were moments of uncertainty—whether lactate’s role was causal or merely correlative, whether SLC5A12 was just a marker or a mediator. It took multiple models, many failed experiments, and an interdisciplinary team to piece this puzzle together.
Looking ahead
Our work opens the door to a new concept: metabolic immune checkpoints. Just as checkpoint inhibitors revolutionised cancer therapy by releasing the brakes on T cells, we believe that targeting metabolic regulators like SLC5A12 could recalibrate immune responses in autoimmune disease.
There’s much still to explore. Do similar mechanisms govern ELS formation in other diseases like lupus or multiple sclerosis? Could SLC5A12 inhibition synergise with existing treatments? And what are the long-term consequences of altering metabolic signalling in immune cells?
These are questions we’re eager to pursue. But for now, we hope this study helps further reframe lactate not as discarded fuel, but as a critical conductor in the symphony of immune organisation.
Dr. Michelangelo Certo is a distinguished researcher at the forefront of inflammation and metabolism. He earned his MSc in Pharmaceutical Chemistry and Technology with honours in 2010, followed by a PhD in Cellular Biochemistry and Pharmacology in 2013, both from the University of Calabria, Italy. His early research at the Centre of Preclinical and Translational Pharmacology focused on unravelling the intricate interplay between the immune system and the brain during ischemic injury, with the goal of identifying groundbreaking immunotherapeutic strategies for stroke.
Recognized for his expertise in Pharmacology by the Department Council at the School of Pharmacy, University of Calabria in 2015, Dr. Certo soon after secured a prestigious Research Fellowship from the Italian Society of Pharmacology. This opportunity led him to University College London, where he expanded his research into the realm of chronic pain, applying his deep understanding of inflammatory pathways to uncover novel therapeutic targets.
Throughout his career, Dr. Certo has been driven by a passion for decoding the inflammatory mechanisms that govern immunity, with a strong emphasis on translational medicine. His work spans cell trafficking, target identification, therapeutic exploitation, and the development of reparative and regenerative therapies. In 2017, he transitioned to Barts and The London School of Medicine and Dentistry at Queen Mary University of London, where he explored the metabolic regulation of immune cell function and the role of key metabolites—such as lactate and fatty acids—as powerful signaling molecules in immune-inflammatory responses.
In 2018, Dr. Certo joined the College of Medicine and Health at the University of Birmingham, where he currently serves as a Senior Research Scientist in Metabolism and Inflammation. His research delves into the complex crosstalk between metabolic and inflammatory pathways, investigating their roles in stroke (funded by the University of Birmingham) and chronic inflammatory diseases, including atherosclerosis (funded by the British Heart Foundation) and Sjögren's syndrome (funded by the Medical Research Council). In addition to his research, he also serves as BMSU Departmental Lead, playing a pivotal role in advancing biomedical research and innovation within the university.
With a career dedicated to bridging fundamental research and therapeutic applications, Dr. Certo continues to push the boundaries of knowledge in inflammation, metabolism, and disease intervention.
Current research project
Dr Michelangelo Certo’s research explores the intricate interplay between metabolic and inflammatory pathways, with a particular focus on how metabolic alterations in inflammation-driven diseases shape immune cell behaviour. These disruptions are not only instrumental in triggering inflammation but also play a key role in sustaining and amplifying it. His work specialises in understanding the metabolic regulation of T cell-mediated immune responses, investigating critical aspects such as cell migration, differentiation, and cytokine production—both under normal physiological conditions and within the context of disease.
A central pillar of his research examines the profound impact of lactate accumulation, which can occur locally in inflamed tissues or systemically during acute and chronic inflammation. His findings have revealed how lactate triggers intracellular metabolic rewiring, significantly influencing immune-inflammatory responses across a range of human diseases. These insights have broad implications for disease progression and the development of targeted therapeutic interventions.
In addition to these areas, Dr Certo investigates the role of lactate in post-stroke inflammation. Ischaemic stroke, caused by an interruption in blood flow to the brain, initiates a cascade of inflammatory responses that can exacerbate neuronal damage and hinder recovery. His research aims to disrupt this cycle by blocking lactate uptake into T cells, thereby reducing the harmful inflammatory burden following a stroke. By targeting lactate transport, his work seeks to mitigate post-stroke inflammation and pave the way for novel therapeutic strategies that improve patient outcomes.
Follow the Topic
-
Nature Metabolism
This journal publishes work from across all fields of metabolism research that significantly advances our understanding of metabolic and homeostatic processes in a cellular or broader physiological context, from fundamental cell biology to basic biomedical and translational research.

Related Collections
With Collections, you can get published faster and increase your visibility.
The expanding therapeutic landscape of GLP 1 receptor agonists
Publishing Model: Hybrid
Deadline: Jan 23, 2027
Microbiome and energy metabolism
Publishing Model: Hybrid
Deadline: Dec 06, 2026


Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in